")
")
| Issue |
Med Sci (Paris)
Volume 27, Number 12, Décembre 2011
|
|
|---|---|---|
| Page(s) | 1045 - 1047 | |
| Section | Les grandes avancées françaises en biologie récompensées par l'Académie des sciences | |
| DOI | https://doi.org/10.1051/medsci/20112712002 | |
| Published online | 23 décembre 2011 | |
Un mécanisme original de perception d’un ion minéral (le nitrate)
Histoire de la découverte chez les plantes et perspectives
An original ion (nitrate) sensing mechanism: story of the discovery in plants and perspectives
Biochimie et physiologie moléculaire des plantes, UMR 5004 CNRS/INRA/SupAgro/UM2, Institut de biologie intégrative des plantes-Claude Grignon, place Viala, 34060 Montpellier Cedex 1, France
* Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Les plantes terrestres prélèvent les ions minéraux nécessaires à leur nutrition essentiellement dans le sol, grâce à leurs racines. Cependant, la disponibilité de ces ions dans le sol varie de manière très importante, à la fois dans le temps et dans l’espace. Pour faire face à ces conditions fluctuantes, les plantes disposent de systèmes d’adaptation très élaborés [1]. En particulier, et contrairement aux animaux, elles sont capables de modifier profondément leur développement post-embryonnaire en fonction des conditions de ressources minérales dans le sol. Un phénomène très intéressant et particulièrement étudié est le fait que les plantes stimulent la croissance de leurs racines spécifiquement là où les nutriments sont les plus abondants, et au contraire la freinent là où ces nutriments font défaut (Figure 1). Ceci est vrai pour un grand nombre d’ions essentiels comme le phosphate (PO42−), le sulfate (SO42−), l’ammonium (NH4+), ou le nitrate (NO3−), et permet ainsi aux végétaux d’optimiser leur nutrition en exploitant préférentiellement les zones du sol riches en minéraux [2].
© 2011 médecine/sciences – Inserm / SRMS
On sait depuis près de 20 ans que cette réponse adaptative met en jeu des systèmes de perception spécifiques des différents ions, permettant aux plantes de les détecter et les localiser dans le milieu. Cependant, les mécanismes moléculaires associés à ces systèmes de perception et impliqués dans les réponses de développement restaient largement inconnus. Notre travail a permis de mettre en évidence le rôle essentiel de la protéine NRT1.1 (un transporteur membranaire de 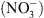 dans le contrôle de la croissance de la plante en fonction de la disponibilité externe en
dans le contrôle de la croissance de la plante en fonction de la disponibilité externe en 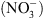 . De manière originale, le mécanisme de perception du
. De manière originale, le mécanisme de perception du 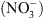 par NRT1.1 met en jeu une particularité fonctionnelle de cette protéine tout à fait inattendue.
par NRT1.1 met en jeu une particularité fonctionnelle de cette protéine tout à fait inattendue.
 |
Figure 1 Illustration de la croissance adaptative des racines des plantes. Quand les racines de plantes rencontrent une zone du sol riche en |
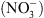
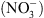
NRT1.1, un transporteur de (NO3−) pas comme les autres
NRT1.1 a été identifiée en 1993 par une approche génétique basée sur un crible de résistance au chlorate [3]. Le chlorate 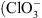 ) est un analogue toxique du
) est un analogue toxique du 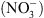 utilisé comme herbicide. Les cribles génétiques chez la plante modèle Arabidopsis thaliana ont permis d’isoler des plantes mutantes résistantes, capables de survivre sur des milieux contenant du
utilisé comme herbicide. Les cribles génétiques chez la plante modèle Arabidopsis thaliana ont permis d’isoler des plantes mutantes résistantes, capables de survivre sur des milieux contenant du 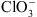 . Certaines de ces plantes se sont avéré être des mutants pour NRT1.1 (appelés mutants chl1) : elles sont donc résistantes au
. Certaines de ces plantes se sont avéré être des mutants pour NRT1.1 (appelés mutants chl1) : elles sont donc résistantes au 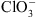 car leurs racines ont un défaut d’absorption de
car leurs racines ont un défaut d’absorption de 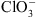 .
.
Si la fonction de transport de 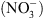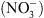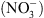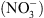 de NRT1.1 a été clairement établie par la suite, il est récemment apparu évident que la perte de cette seule fonction de transport de
de NRT1.1 a été clairement établie par la suite, il est récemment apparu évident que la perte de cette seule fonction de transport de 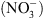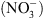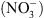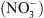 ne pouvait expliquer un certain nombre de désordres physiologiques ou développementaux affectant les mutants chl1 [4].
ne pouvait expliquer un certain nombre de désordres physiologiques ou développementaux affectant les mutants chl1 [4].
En particulier, les mutants chl1 ont perdu une partie des réponses adaptatives déclenchées par les systèmes de perception du 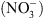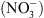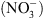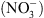 , notamment celle correspondant à la croissance racinaire préférentielle dans les zones du milieu riches en cet ion (Figure 1) [5]. Ceci pouvait suggérer que NRT1.1 possède en fait une double fonction, non seulement de transporteur mais aussi de senseur de
, notamment celle correspondant à la croissance racinaire préférentielle dans les zones du milieu riches en cet ion (Figure 1) [5]. Ceci pouvait suggérer que NRT1.1 possède en fait une double fonction, non seulement de transporteur mais aussi de senseur de 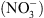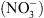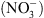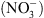 (voir [4] pour le détail de cette hypothèse).
(voir [4] pour le détail de cette hypothèse).
Cependant, il fallait découvrir les mécanismes de cette possible fonction de senseur et de son effet sur le développement racinaire.
L’hypothèse : NRT1.1 contrôle le transport d’une hormone végétale, l’auxine
En plus des phénotypes inexpliqués des mutants chl1, deux éléments d’information ont permis de diriger les recherches pour élucider le mécanisme de signalisation dépendant de NRT1.1.
Premièrement, NRT1.1 appartient à une famille de transporteurs (nommée PTR pour peptide transporter), dont la spécificité de substrat n’est pas stricte. Par exemple, les plus proches homologues caractérisés de NRT1.1 sont : (1) une protéine du colza (Brassica napus), BnNRT1.2, capable de transporter aussi bien des acides aminés comme l’histidine ou la lysine que du 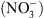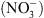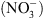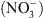 ; (2) une protéine de l’aulne glutineux1 (Alnus glutinosa), capable de transporter différents acides organiques comme le malate ou le succinate. Ceci suggérait que NRT1.1 puisse également transporter d’autres solutés que du
; (2) une protéine de l’aulne glutineux1 (Alnus glutinosa), capable de transporter différents acides organiques comme le malate ou le succinate. Ceci suggérait que NRT1.1 puisse également transporter d’autres solutés que du 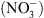 , notamment des acides organiques ou aminés.
, notamment des acides organiques ou aminés.
Deuxièmement, un acide organique particulier, structurellement proche de l’acide aminé tryptophane, est l’acide indole acétique (AIA), hormone végétale connue sous le nom d’auxine. L’auxine est un puissant morphogène qui joue un rôle central dans le contrôle du développement et de la différenciation cellulaire, notamment au niveau des racines latérales. En effet, le développement des primordia de racines latérales nécessite l’établissement et le maintien d’un gradient de concentration d’auxine (avec un maximum à l’apex des primordia) [6]. Dans les racines, comme dans les feuilles, ce gradient local dépend de l’activité de transporteurs spécifiques, qui canalisent l’hormone vers l’apex du primordium, ou au contraire l’en éloignent [6].
Ces considérations, et la constatation que NRT1.1 contrôle la croissance des racines latérales en réponse au 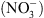 [5], nous ont tout naturellement conduit à suggérer que NRT1.1 pourrait être un transporteur d’auxine. Cette hypothèse a été testée en système hétérologue (ovocytes de xénope, levure) ainsi que dans la plante. Dans tous les cas, nous avons montré que NRT1.1 facilite effectivement le transport de l’auxine. Mais cela ne suffisait pas à expliquer le lien avec le rôle de perception du
[5], nous ont tout naturellement conduit à suggérer que NRT1.1 pourrait être un transporteur d’auxine. Cette hypothèse a été testée en système hétérologue (ovocytes de xénope, levure) ainsi que dans la plante. Dans tous les cas, nous avons montré que NRT1.1 facilite effectivement le transport de l’auxine. Mais cela ne suffisait pas à expliquer le lien avec le rôle de perception du 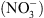 joué par cette protéine. Ce n’est que lors des mesures de transport d’auxine en présence de
joué par cette protéine. Ce n’est que lors des mesures de transport d’auxine en présence de 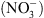 (les deux substrats connus de NRT1.1) que nous avons observé que le
(les deux substrats connus de NRT1.1) que nous avons observé que le 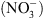 inhibe le transport d’auxine assuré par NRT1.1 (Figure 2A).
inhibe le transport d’auxine assuré par NRT1.1 (Figure 2A).
 |
Figure 2 Rôle de la protéine NRT1.1 dans le contrôle de l’homéostasie de l’auxine et de la croissance des racines latérales. A. Au niveau cellulaire, NRT1.1 facilite le transport de l’auxine. Ce transport est inhibé par les fortes concentrations de |
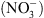
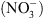
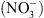
Nous avons donc proposé le schéma suivant : NRT1.1 contrôle la croissance racinaire en réponse au 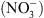 parce que son activité de transport d’auxine module l’homéostasie de l’hormone en fonction de la concentration externe en
parce que son activité de transport d’auxine module l’homéostasie de l’hormone en fonction de la concentration externe en 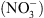 . Ce modèle été validé in planta, par la double analyse : (1) de la localisation tissulaire de NRT1.1, qui a permis de montrer que ce transporteur est spécifiquement exprimé dans les territoires cellulaires assurant le reflux de l’auxine hors de la racine latérale vers la racine primaire (Figure 2B, flèches jaunes), et (2) des gradients locaux d’auxine dans les racines latérales chez les plantes sauvages et les mutants chl1. L’analyse de ces derniers a en effet révélé que la faible croissance racinaire en l’absence de
. Ce modèle été validé in planta, par la double analyse : (1) de la localisation tissulaire de NRT1.1, qui a permis de montrer que ce transporteur est spécifiquement exprimé dans les territoires cellulaires assurant le reflux de l’auxine hors de la racine latérale vers la racine primaire (Figure 2B, flèches jaunes), et (2) des gradients locaux d’auxine dans les racines latérales chez les plantes sauvages et les mutants chl1. L’analyse de ces derniers a en effet révélé que la faible croissance racinaire en l’absence de 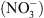 s’accompagne d’un défaut d’accumulation de l’hormone à l’apex des racines, et que ce défaut d’accumulation est dû à l’absence de NRT1.1 (l’auxine s’accumule fortement en l’absence de
s’accompagne d’un défaut d’accumulation de l’hormone à l’apex des racines, et que ce défaut d’accumulation est dû à l’absence de NRT1.1 (l’auxine s’accumule fortement en l’absence de 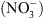 chez les mutants chl1).
chez les mutants chl1).
Le modèle de perception du (NO3−) et du contrôle de la croissance de la racine latérale
Ainsi, le transporteur NRT1.1 est impliqué dans le couplage biophysique des signalisations auxine et 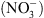 . En d’autres termes, NRT1.1 transforme les variations de disponibilité externe en
. En d’autres termes, NRT1.1 transforme les variations de disponibilité externe en 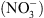 en une modification des flux d’auxine au sein de l’organe.
en une modification des flux d’auxine au sein de l’organe.
Le modèle de développement qui en découle est le suivant : dans des situations de faible disponibilité externe en 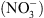 , NRT1.1 se comporte comme un transporteur d’auxine qui exporte l’hormone hors de la racine latérale, empêchant ainsi son accumulation à l’apex (Figure 2B) ce qui inhibe la croissance. À l’inverse, lorsque le
, NRT1.1 se comporte comme un transporteur d’auxine qui exporte l’hormone hors de la racine latérale, empêchant ainsi son accumulation à l’apex (Figure 2B) ce qui inhibe la croissance. À l’inverse, lorsque le 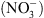 est localement abondant, il réprime le transport d’auxine médié par NRT1.1, ce qui a pour conséquence de favoriser l’accumulation d’auxine dans les racines latérales, et donc de stimuler la croissance de ces racines dans les milieux riches en
est localement abondant, il réprime le transport d’auxine médié par NRT1.1, ce qui a pour conséquence de favoriser l’accumulation d’auxine dans les racines latérales, et donc de stimuler la croissance de ces racines dans les milieux riches en 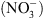 (Figure 2B). Récemment, des résultats d’une équipe japonaise suggèreraient que ce mécanisme permet aussi de contrôler la croissance des parties aériennes [7]. L’ensemble de notre travail a été valorisé dans un article dans la revue Developmental Cell [8].
(Figure 2B). Récemment, des résultats d’une équipe japonaise suggèreraient que ce mécanisme permet aussi de contrôler la croissance des parties aériennes [7]. L’ensemble de notre travail a été valorisé dans un article dans la revue Developmental Cell [8].
Perspectives
Ce mécanisme d’interaction entre les transports membranaires d’une hormone et d’un nutriment minéral est à notre connaissance le premier décrit. Il ouvre des perspectives en agriculture car la gestion optimisée de la croissance et de l’architecture du système racinaire des plantes cultivées est un objectif essentiel, encore peu pris en compte pour l’amélioration variétale [9].
Par ailleurs, des protéines homologues à NRT1.1 ont été identifiées chez d’autres organismes, et chez les animaux en particulier. Du fait des proximités structurales et fonctionnelles de l’auxine et de la sérotonine [10], il n’est donc pas impossible que la découverte de ce mécanisme puisse être transposée au domaine animal, et ait à long terme des conséquences en médecine.
Conflit d’intérêts
Les auteurs déclarent n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article.
L’aulne, parfois appelé « aune », ou « verne », est l’arbre typique des bords de cours d’eau et des zones marécageuses du fait de ses exigences écologiques. Le bois d’aulne est imputrescible, qualité qui lui a valu de beaucoup servir auparavant comme matériau de construction en contact avec l’eau : conduites, pilotis, soubassement… Venise est bâtie sur des pieux d’aulne !
Références
- Krouk G, Crawford NM, Coruzzi GM, Tsay YF. Nitrate signaling: adaptation to fluctuating environments. Curr Opin Plant Biol 2010 ; 13 : 266–273. [CrossRef] [PubMed] [Google Scholar]
- Drew MC. Comparison of the effects of a localized supply of phosphate, nitrate, ammonium and potassium on the growth of the seminal root system, and the shoot, in barley. New Phytol 1975 ; 75 : 479–490. [CrossRef] [Google Scholar]
- Tsay YF, Schroeder JI, Feldmann KA, Crawford NM. The herbicide sensitivity gene CHL1 of Arabidopsis encodes a nitrate-inducible nitrate transporter. Cell 1993 ; 72 : 705–713. [CrossRef] [PubMed] [Google Scholar]
- Gojon A, Krouk G, Perrine-Walker F, Laugier E. Nitrate transceptor(s) in plants. J Exp Bot 2011 ; 62 : 2299–2308. [CrossRef] [PubMed] [Google Scholar]
- Remans T, Nacry P, Pervent M, et al. The Arabidopsis NRT1.1 transporter participates in the signaling pathway triggering root colonization of nitrate-rich patches. Proc Natl Acad Sci USA 2006 ; 103 : 19206–19211. [CrossRef] [Google Scholar]
- Benkova E, Michniewicz M, Sauer M, et al. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003 ; 115 : 591–602. [CrossRef] [PubMed] [Google Scholar]
- Hachiya T, Mizokami Y, Miyata K, et al. Evidence for a nitrate-independent function of the nitrate sensor NRT1.1 in Arabidopsis thalianaJ Plant Res 2011 ; 124 : 425–430. [CrossRef] [PubMed] [Google Scholar]
- Krouk G, Lacombe B, Bielach A, et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev Cell 2010 ; 18 : 927–937. [Google Scholar]
- Herder GD, Van Isterdael G, Beeckman T, De Smet I. The roots of a new green revolution. Trends Plant Sci 2010 ; 15 : 600–607. [CrossRef] [PubMed] [Google Scholar]
- Pelagio-Flores R, Ortiz-Castro R, Mendez-Bravo A, et al. Serotonin, a tryptophan-derived signal conserved in plants and animals, regulates root system architecture probably acting as a natural auxin inhibitor in Arabidopsis thaliana. Plant Cell Physiol 2011 ; 52 : 490–508. [Google Scholar]
Liste des figures
|
Figure 1 Illustration de la croissance adaptative des racines des plantes. Quand les racines de plantes rencontrent une zone du sol riche en |
| Dans le texte | |
|
Figure 2 Rôle de la protéine NRT1.1 dans le contrôle de l’homéostasie de l’auxine et de la croissance des racines latérales. A. Au niveau cellulaire, NRT1.1 facilite le transport de l’auxine. Ce transport est inhibé par les fortes concentrations de |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.