")
")
| Issue |
Med Sci (Paris)
Volume 25, Number 10, Octobre 2009
|
|
|---|---|---|
| Page(s) | 802 - 803 | |
| Section | Flash | |
| DOI | https://doi.org/10.1051/medsci/20092510802 | |
| Published online | 15 octobre 2009 | |
Naissance de Mito et Tracker
Prélude à la correction des mutations de l’ADN mitochondrial chez l’homme ?
Birth of Mito and Tracker: mitochondrial gene replacement
1
Médecine/Sciences et École normale supérieure, Paris, France
2
Médecine/Sciences et Inserm U935, Hôpital Paul Brousse, 94817 Villejuif Cedex, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Quelquefois, une centrale d’énergie devient une bombe à retardement… L’ADN mitochondrial (ADNmt), hérité de la mère compte tenu de l’apport cytoplasmique négligeable du spermatozoïde, est 10 fois plus susceptible aux mutations que l’ADN nucléaire, peut-être par son exposition directe à une forte concentration de radicaux libres, à l’absence d’histones ou à la moindre efficacité des mécanismes de réparation. Plus de 150 mutations de l’ADNmt ont été répertoriées, transmissibles aux générations suivantes, responsables de maladies sévères pour lesquelles il n’y a pas de traitement et qui sont exceptionnellement détectables par une stratégie de diagnostic préimplantatoire [1, 2]. Peut-on agir en amont et empêcher la transmission via l’ovocyte des milliers de molécules d’ADNmt défectueuses ? En théorie, on pourrait diluer les mitochondries mutantes en transférant du cytoplasme normal d’un ovocyte immature, ou transférer dans un ovocyte sain l’ADN nucléaire sous forme de pronucléus ou de vésicule germinale (selon que l’on est après ou avant fécondation), toutes approches testées : mais aucune ne corrige suffisamment l’hétéroplasmie (la répartition des mitochondries rend inévitable la contamination dans le cas du transfert de vésicule germinale) et elles peuvent entraîner des anomalies chromosomiques inacceptables.
 |
Figure 1. Stratégie de correction de mutations de l’ADN mitochondrial par transfert exclusif de l’ADN nucléaire chez le macaque. L’originalité de la statégie consiste à transférer le complexe associant le fuseau mitotique et les chromosomes (caryoplaste), complexe prélevé dans un ovocyte mature arrêté au stade métaphase II de la méiose (A). Le complexe ne contient pratiquement pas de mitochondries. Il est fusionné à un ovocyte receveur au même stade de la méiose, et qui a été énucléé au préalable (par retrait du complexe caryoplaste) (B). Après intégration du complexe, la fécondation est provoquée par ICSI (intracytoplasmic sperm injection), ce qui déclenche les premières divisions embryonnaires. Les embyrons ainsi obtenus, peuvent être soit utilisés pour dériver des lignées de cellules ES, soit réimplantés dans l’utérus de femelles macaques (C). Sur 9 femelles ayant reçu 15 blastocystes, 3 grossesses ont été obtenues, aboutissant à la naissance de 4 macaques nouveau-nés sains (une grossesse géméllaire) dont l’ADN mitochondrial a pour origine l’ovocyte receveur, et le patrimoine génétique nucléaire l’ovocyte donneur. |
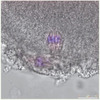 |
Figure 2. Transfert du caryoplaste dans un ovocyte. Cette photo, comme celle de la couverture, nous a été aimablement donnée par le laboratoire de S. Mitalipov. |
L’équipe de S. Mitalipov (centre de primatologie de l’Orégon, États-Unis) publie dans Nature une nouvelle approche extrêmement élégante de transfert exclusif de l’ADN nucléaire, et fait avec succès la preuve de concept de son application chez le macaque [3]. La stratégie consiste à transférer, dans un ovocyte receveur mature bloqué en métaphase II et énucléé, l’ADN nucléaire (sous la forme du complexe chromosomes-fuseau mitotique pauvre en mitochondries) prélevé d’un ovocyte « donneur » (et qui, dans le cadre d’une application thérapeutique, pourrait être porteur de mutations de l’ADNmt). On pensait impossible un tel transfert d’ADN nucléaire entre ovocytes à ce stade avancé de maturation en raison de la difficulté de visualisation et d’isolement du fuseau métaphasique une fois la membrane nucléaire rompue, et de la vulnérabilité des chromosomes dépourvus d’enveloppe nucléaire. L’équipe, qui a une longue expérience du transfert nucléaire somatique chez les primates (qui implique l’énucléation de l’ovocyte) et a dérivé avec succès des lignées de cellules souches embryonnaires par cette technique [4], a surmonté ces obstacles en utilisant des technologies innovantes de visualisation non invasive du complexe fuseau-chromomoses par microscopie polarisée, ne nécessitant ni colorant ni rayons UV, ce qui évite d’endommager l’ADN. Le fuseau mitotique est dépourvu de mitochondries et le faible volume du caryoplaste (désigne l’ensemble fuseau et cytoplasme environnant) - correspondant à 1,5 % de l’oocyte - minimise la contamination par l’ADNmt du donneur. Cette contamination est au contraire abondante dans l’espace périnucléaire qui accompagne la vésicule germinale ou le pronucléus entier lorsque ceux-ci sont transférés dans des ovocytes sains énucléés, ce qui, en outre, perturbe la maturation ultérieure de l’ovocyte. Un élément critique du travail était le choix de l’agent fusogène, ici un extrait du virus de Sendai qui induit la fusion du caryoplaste et du cytoplasme de l’ovocyte receveur énucléé sans rompre le blocage en métaphase de la méiose II, indispensable à la fécondation ultérieure. La difficulté réside aussi dans la parfaite synchronie qui est requise dans les phases de la méiose des deux ovocytes donneur et receveur. Après fécondation par injection intracytoplasmique (ICSI) de sperme, des blastocystes se forment normalement, dont on peut dériver des lignées de cellules souches embryonnaires (ES) pluripotentes typiques. Surtout, 15 de ces blastocystes ont été implantés dans les oviductes de 9 femelles : il en est résulté 3 grossesses (proportion identique à celle obtenue par ICSI) aboutissant à la naissance de 4 petits macaques sains dont des jumeaux. La recherche très soigneuse d’une hétéroplasmie, notamment par séquençage des polymorphismes de la région hypervariable 1 de la boucle D de l’ADN mitochondrial (rhDHV1) dans les tissus de trois petits macaques (seuls trois étaient nés lors de l’acceptation de l’article) et dans les cellules ES, a été négative : le taux de contamination détectable de la préparation fuseau-chromosomes par de l’ADNmt du donneur se situe donc en dessous du seuil de sensibilité (3 %) de différentes méthodes de détection (séquençage, qPCR). Quelle faisabilité chez l’homme ? Outre la discussion éthique qu’implique la réimplantation d’embryons issus de gamètes modifiés (ce qu’interdit l’actuelle réglementation française), il est trop tôt pour juger pleinement de la sécurité et de l’efficacité de cette technique. Notamment, même si aucune séquence du génome viral n’a été détectée, il serait sage de substituer l’extrait du virus de Sendai par un agent non viral ; seconde question : la possibilité à long terme d’une dérive génétique et/ou d’un biais de distribution des mitochondries pendant la division cellulaire, entraînant une augmentation de l’ADNmt du donneur et une hétéroplasmie dans certains tissus ou dans les générations suivantes. Mito et Tracker, qui seront suivis de très près, nous le diront… Mais quel superbe travail !
Conflit d’intérêts
Les auteurs déclarent n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article.
Références
- Taylor RW, Turnbull DM. Mitochondrial DNA mutations in human disease. Nat Rev Genet 2005; 6 : 389–402. [Google Scholar]
- Rustin P, T Jacobs H, Dietrich A, et al. Adresser du matériel allogène dans le compartiment mitochondrial : un défi pour comprendre la physiologie mitochondriale et une perspective pour la thérapie. Med Sci (Paris) 2007; 23 : 519–25. [Google Scholar]
- Tachibana M, Sparman M, Sritanaudomchai H, et al. Mitochondrial gene replacement in primate offspring and embryonic stem cells. Nature 2009; 26 août (online). [Google Scholar]
- Byrne JA, Pedersen DA, Clepper LL, et al. Producing primate embryonic stem cells by somatic cell nuclear transfer. Nature 2007; 450 : 497–502. [Google Scholar]
© 2009 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Stratégie de correction de mutations de l’ADN mitochondrial par transfert exclusif de l’ADN nucléaire chez le macaque. L’originalité de la statégie consiste à transférer le complexe associant le fuseau mitotique et les chromosomes (caryoplaste), complexe prélevé dans un ovocyte mature arrêté au stade métaphase II de la méiose (A). Le complexe ne contient pratiquement pas de mitochondries. Il est fusionné à un ovocyte receveur au même stade de la méiose, et qui a été énucléé au préalable (par retrait du complexe caryoplaste) (B). Après intégration du complexe, la fécondation est provoquée par ICSI (intracytoplasmic sperm injection), ce qui déclenche les premières divisions embryonnaires. Les embyrons ainsi obtenus, peuvent être soit utilisés pour dériver des lignées de cellules ES, soit réimplantés dans l’utérus de femelles macaques (C). Sur 9 femelles ayant reçu 15 blastocystes, 3 grossesses ont été obtenues, aboutissant à la naissance de 4 macaques nouveau-nés sains (une grossesse géméllaire) dont l’ADN mitochondrial a pour origine l’ovocyte receveur, et le patrimoine génétique nucléaire l’ovocyte donneur. |
| Dans le texte | |
|
Figure 2. Transfert du caryoplaste dans un ovocyte. Cette photo, comme celle de la couverture, nous a été aimablement donnée par le laboratoire de S. Mitalipov. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.