")
")
| Issue |
Med Sci (Paris)
Volume 25, Number 4, Avril 2009
|
|
|---|---|---|
| Page(s) | 337 - 340 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2009254337 | |
| Published online | 15 avril 2009 | |
Maladies neurodégénératives et diabète
Glucose homeostasis, diabetes and neurodegenerative disorders
1
Inserm U679, Hôpital Pitié-Salpêtrière, Paris, France. Adresse actuelle : Université Montpellier 2, Laboratoire MMDN, Inserm U710, Bâtiment 24, CC105, Place Eugène Bataillon, 34095 Montpellier Cedex 5, France
2
Laboratoire d’Endocrinologie Moléculaire, IURC (UMR 204), Université Montpellier I, Montpellier, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Compte tenu du vieillissement continu de la population, et de l’augmentation de la prévalence du diabète sucré et des maladies neurodégénératives, on peut s’interroger sur une relation possible entre l’action de l’insuline ou des IGF (insulin-like growth factor) et le déterminisme des maladies neurodégénératives dont les représentants majeurs sont la maladie d’Alzheimer (MA) et la maladie de Parkinson (MP). Les interactions entre l’insuline et le système nerveux ont été ignorées pendant de nombreuses années. Ce n’est que grâce aux expériences récentes d’invalidation - dans les tissus nerveux - des gènes codant pour les récepteurs de l’insuline (RI) et de l’IGF-1 (R-IGF-1) que la conception classique d’un cerveau insensible à l’insuline a été remise en question [1].
Expression de l’insuline, des IGF et de leurs récepteurs dans le système nerveux
De nombreuses observations se sont accumulées montrant que les RI s’expriment dans les cellules neuronales et gliales de plusieurs régions du système nerveux central - les bulbes olfactifs, l’hippocampe, l’hypothalamus, la substantia nigra et le cortex [2, 3]. Si les effets métaboliques sur le cerveau sont moins importants, le transport du glucose dépendant également de la distribution régionale des transporteurs GLUT4 et GLUT8, les effets de l’insuline sur la mémoire, l’apprentissage et le comportement alimentaire ont reçu une attention particulière ces dernières années [2].
Les mécanismes impliqués dans ces effets demeurent complexes. Par exemple, au niveau de l’hypothalamus, l’insuline serait capable de réguler la sécrétion des neuropeptides NPY, POMC (pro-opiomélanocortine), AgRP (agoutirelated protein) et CART (cocaineamphetamine-related transcript) en synergie avec les effets de la leptine [3]. Au niveau post-synaptique, elle modulerait l’action des neurotransmetteurs (acétylcholine et noradrénaline) et le recrutement des récepteurs de type GABAA [3]. Cependant, le mécanisme le plus intéressant concerne les effets de l’insuline et des IGF sur la différenciation, la survie et la régénération neuronales [4]. En effet, in vitro, les IGF stimulent la croissance des neurites de cellules de neuroblastome. Les IGF participent également à la régénération des neurones moteurs ou sensitifs. Lors d’une lésion expérimentale (pression sur le nerf sciatique par exemple), la régénération axonale qui en résulte s’accompagne d’une surexpression des IGF dans les cellules de Schwann. Le fait que l’application locale d’IGF-1 ou d’IGF-2 améliore la régénération des nerfs sensitifs suggère le rôle physiologique de ces molécules. Cela concorde avec l’observation que les cellules neuronales ainsi que les axones et les terminaisons nerveuses expriment les récepteurs des IGF et que ces hormones peuvent pénétrer dans l’espace péri-neuronal par les capillaires fenestrés [4]. Les effets des IGF et de l’insuline s’entrecroisent au niveau cellulaire, grâce notamment à leurs homologies de structure. En effet, les RI et les R-IGF appartiennent à une famille de protéines homologues possédant une activité tyrosine kinase. L’IGF-1 agit via un récepteur spécifique, mais celui-ci peut également être activé par l’IGF-2 ou l’insuline. De plus, les RI et les R-IGF-1 forment à la surface membranaire des molécules hybrides. Cette intrication peut avoir une importance fonctionnelle, car des taux élevés d’insuline, comme c’est notamment le cas lors des phases précoces du diabète du type 2, pourraient stimuler les récepteurs des IGF.
Relation entre diabète et maladies neurodégénératives
La relation entre le diabète et les maladies neurodégénératives impose une analyse plus approfondie de la résistance à l’insuline et de sa pathogénie dans les maladies génétiques ou les maladies neurodégénératives les plus fréquentes. Dans les syndromes génétiques de résistance sévère à l’insuline due à des mutations des gènes de la voie de l’insuline ou de l’IGF-1 [5], l’atteinte du système nerveux (retard mental ou altération du comportement) est bien présente, sans que l’on puisse affirmer une relation directe entre les phénomènes neurodégénératifs et métaboliques (Tableau 1). Cependant, un certain degré de résistance à l’insuline accompagne des maladies génétiques ayant une composante neurodégénérative, comme l’ataxie télangiectasie, les syndromes de Werner ou de Prader-Willy [6].
Maladies associées à des mutations pathogènes des gènes impliqués dans la cascade de signalisation de l’insuline et de l’IGF-1. (http://www.orphanet.com). a : récepteur de l’insuline ; b : récepteur de l’IGF-1 ; c : growth factor receptor-bound protein 10 ; d : Grb-10 interacting GYF protein 2 ; e : hypoglycémie à jeun ; f : retard de croissance intra-utérin ; ND : non déterminé lors de nos travaux détaillés dans [10].
IDE (insulin degrading enzyme) et maladie d’Alzheimer
La relation entre la résistance à l’insuline et les deux maladies neurodégénératives les plus fréquentes est bien étudiée [3]. Dans la maladie d’Alzheimer, les études épidémiologiques ont montré l’effet synergique du diabète et du génotype ε4 de l’APOE (apolipoprotéine E) dans l’augmentation du risque de développer cette affection [7, 15], ainsi qu’une relation entre la maladie d’Alzheimer et la résistance à l’insuline. Des arguments cliniques ont été apportés par l’équipe de S. Craft à Seattle (États-Unis) montrant l’effet bénéfique de l’administration d’insuline sur la mémoire chez des adultes sains ou affectés par la maladie d’Alzheimer. Ces effets nécessitent des doses d’insuline plus élevées chez les malades Alzheimer que chez les adultes sains [2]. De plus, de nombreuses études in vitro ont montré que l’insuline intervenait dans la libération intracellulaire d’amyloïde β (Aβ), l’accélération du processus de transport vers la membrane cellulaire et l’agrégation des fragments protéiques issus de l’APP (β-amyloid precursor protein). L’accumulation progressive de dépôts de peptide β-amyloïde (βA) dans le cerveau représente une caractéristique des patients atteints de la maladie d’Alzheimer [16]. Le taux d’Aβ est la résultante de la synthèse mais aussi de la dégradation protéolytique du peptide [16]. La relation entre l’insuline et la concentration en Aβ est complexe : ainsi, l’une des enzymes impliquées dans la dégradation de l’Aβ est IDE (insulin degrading enzyme), une thio-métalloprotéinase qui s’exprime dans le cerveau et participe à la clairance de l’Aβ du tissu cérébral. Si l’enzyme clive l’insuline in vitro, son rôle physiologique dans le catabolisme de l’insuline in vivo est encore incertain. Une faible activité de cette IDE se traduirait par l’accumulation d’une part de l’Aβ et d’autre part de l’insuline (hyperinsulinisme). Par exemple, le rat GK (Goto et Kakisaki) qui est un modèle classique de diabète de type 2 non obèse, possède deux mutations pathogènes dans le gène Ide [8]. Lorsque les séquences Ide mutées sont exprimées dans des cellules en culture, on observe une réduction de 31 % de la dégradation de l’insuline. La délétion homozygote de ce même gène chez la souris Ide−/− s’accompagne d’une diminution de 50 % du taux de peptide Aβ dans le cerveau, et ce résultat est dupliqué dans des cultures primaires de neurones Ide−/− [9]. Chez l’homme, le gène Ide est localisé sur le chromosome 10 (10q23-q25) au niveau d’un locus également en cause dans le diabète sucré (OMIM #146680). Ces données suggèrent qu’une des explications de l’association entre diabète de type 2 et maladie d’Alzheimer avec accumulation de peptide Aβ serait un défaut de l’IDE et, dans ce cas, l’hyperinsulinisme serait l’altération primaire et la résistance à l’insuline une conséquence secondaire.
Insuline et maladie de Parkinson
La possibilité d’une relation entre maladie de Parkinson et résistance à l’insuline a elle aussi été bien étudiée. Quelques études épidémiologiques ont montré que le diabète est un facteur de risque majeur qui accélère la détérioration de la fonction motrice ou l’état cognitif [10]. Les patients atteints de maladie de Parkinson ont une résistance à l’insuline et, dans 50 à 80 % des cas, une intolérance glucidique. La L-DOPA et la bromocriptine utilisées dans le traitement de la maladie de Parkinson influencent aussi l’homéostasie glucidique, le premier produit étant hyperglycémiant et le second insulino-sensibilisant. Mais, plus important, l’examen neuropathologique du cerveau de sujets atteints de maladie de Parkinson a révèlé une diminution de l’expression des RI ou de la tyrosine hydroxylase dans la substantia nigra. La relation entre ces deux maladies neurodégénératives et l’action de l’insuline et des IGF peut donc résulter de l’atteinte de gènes de susceptibilité communs aux maladies neurodégénératives et métaboliques. Elle peut aussi témoigner de la connexion au niveau moléculaire de voies de signalisation de facteurs communs à ces deux processus.
Croisement entre voies de signalisation de l’insuline et de l’homéostasie neuronale
La croisement entre l’action des IGF et la maladie d’Alzheimer peut se faire aussi au niveau de la cascade de signalisation. Par exemple, des isoformes de la sousunité p85α de la PI3K (phosphoinositide 3-kinase), qui joue un rôle crucial dans la signalisation de l’insuline, sont associées à un risque accru de maladie d’Alzheimer [3]. D’autre part, toute une série de protéines intervenant dans les voies de signalisation insuline/IGF ont un rôle potentiel dans l’homéostasie neuronale. Par exemple, la voie PKB (ou Akt), acteur important en aval du récepteur de l’insuline aurait un rôle neuroprotecteur via la protéine pro-apoptotique Bad, le facteur CREB (cAMP response elementbinding) ou encore le facteur de transcription FOXO-1 (forkhead transcription factor), qui contrôle, entre autres, la transcription des protéines de la famille Bcl-2. Plus en aval, la GSK-3 (glycogen synthase kinase 3) aurait un rôle dans la phosphorylation des substrats comme CRMP2 (collapsin response mediator proteins), la préseniline-1 (les présénilines contrôlent la maturation de la βAPP, et les mutations de ces protéines accélèrent le processus neurodégénératif de la maladie d’Alzheimer), la protéine Tau et même le précurseur d’amyloïde [3, 4]. Il semblerait donc que ces acteurs de la signalisation soient des intermédiaires communs de l’action des facteurs de croissance neuronale (NGF, nerve growth factor ; BDNF, brain-derived growth factor ; et NT (neurotrophin) 3, -4 et -5) et, par ce biais, représentent des candidats potentiels des défauts moléculaires des maladies neurodégératives (Figure 1).
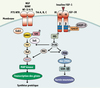 |
Figure 1. Les voies de signalisation de l’insuline et de l’IGF-1 et des facteurs de croissance neuronale. |
Cette dernière hypothèse a été récemment confortée par nos observations - dans le laboratoire de R.J. Smith aux Etats-Unis - du rôle de la protéine GIGYF2 (Grb10-interacting GYF protein-2) dans la maladie de Parkinson (Tableau 1). Cette protéine, impliquée dans la voie de signalisation de l’IGF-1, se lie par son domaine GYF à la protéine adaptatrice Grb10, ce complexe interagissant avec les récepteurs IGF-1 via le domaine SH2 de Grb10 (Figure 1). La résultante de cette interaction est l’inhibition de la phosphorylation sur tyrosine des substrats IRS (insulin responsive substrate), mais également le blocage concomitant de l’accès aux phosphatases, mettant ainsi les récepteurs dans un état actif latent [11–13]. L’implication de ce mécanisme dans la pathologie reste inconnue. Cependant, le rôle spécifique de la protéine adaptatrice Grb10 dans le développement neurologique a déjà été suggéré par la découverte de mutations pathogènes dans le syndrome de Silver-Russell, qui associe un retard de croissance intra-utérin et un retard du développement psychomoteur ou du langage [13]. Notre étude du gène GIGYF2 par séquençage direct dans 249 cas familiaux (italiens et français) de la maladie de Parkinson et 227 témoins nous a permis d’identifier une mutation (faux-sens) du gène GIGYF2 chez 6 % des sujets atteints de maladie de Parkinson [14]. D’autre part, nous avons pu observer une importante variabilité de type insertion et délétion de codons glutamine (Glu). Or, il a déjà été montré que l’expansion polyglutamine était responsable de la maladie d’Huntington. D’autres part, la protéine GYGF2 s’exprime dans le cerveau et présente donc un intérêt particulier, puisque pour la première fois des altérations du gène codant pour une protéine intervenant dans la voie de signalisation de l’IGF-1 sont associées directement à une maladie neuro-dégénérative. L’étude approfondie de l’impact de ces variations géniques sur la fonction de la protéine nous apportera de précieuses informations sur son mode d’action et sur le croisement des voies métaboliques et neurologiques.

Remerciements
Nous souhaitons remercier le Professeur Alexis Brice qui nous a encouragés dans la rédaction de cet article.
Références
- Plum L, Belgardt BF, Bruning JC. Central insulin action in energy and glucose homeostasis. J Clin Invest 2006; 116 : 1761–6. [Google Scholar]
- Zhao WQ, Alkon DL. Role of insulin receptor in learning and memory. Mol Cell Endocrinol 2001; 177 : 125–34. [Google Scholar]
- Craft S, Watson GS. Insulin and neurodegenerative disease: shared and specific mechanisms. Lancet Neurol 2004; 3 : 169–78. [Google Scholar]
- Yasuda H, Terada M, Maeda K, et al. Diabetic neuropathy and nerve regenaration. Prog Neurobiol 2003; 69 : 229–85. [Google Scholar]
- Abuzzahab MJ, Schneider A, Goddard A, et al. Igf-I receptor mutations resulting in intrauterine and postnatal growth retardation. N Engl J Med 2003; 349 : 2211–22. [Google Scholar]
- Ristow M. Neurodegenerative disorders associated with diabetes mellitus. J Mol Med 2004; 82 : 510–29. [Google Scholar]
- Peila R, Rodriguez BI, Launer LJ. Type 2 diabetes, APOE gene, and the risk for dementia and related pathologies: The Honolulu-Asia aging study. Diabetes 2002; 51 : 1256–62. [Google Scholar]
- Qiu WQ, Walsh DM, Ye Z, et al. Insulin degrading enzyme regulates extracellular levels of amyloid beta-protein by degradation. J Biol Chem 1998; 273 : 32730–8. [Google Scholar]
- Farris W, Mansourian S, Chang Y, et al. Insulin degrading enzyme regulates the levels of insulin, amyloid β-protein, and β-amyloid precursor protein intracellular domain in vivo. Proc Natl Acad Sci USA 2003; 100 : 4162–7. [Google Scholar]
- Hu G, Jousilahti P, Bidel S, et al. Type 2 diabetes and the risk of Parkinson’s disease. Diabetes Care 2007; 30 : 842–7. [Google Scholar]
- Dufresne AM, Smith RJ. The adapter protein Grb10 is an endogenous negative regulator of insulin-like growth factor signaling. Endocrinology 2005; 146 : 4399–409. [Google Scholar]
- Giovannone B, Lee E, Laviola L, et al. Two novel proteins that are linked to insulin-like growth factor (Igf-I) receptors by the Grb10 adapter and modulate Igf-I signaling. J Biol Chem 2003; 278 : 31564–73. [Google Scholar]
- Yoshihashi H, Maeyama K, Kosaki R, et al. Imprinting of human Grb10 and its mutations in two patients with Russell-Silver syndrome. Am J Hum Genet 2000; 67 : 476–82. [Google Scholar]
- Lautier C, Goldwurm S, Durr A, et al. Mutations in the Gigyf2 (Tnrc15) gene at the Park11 locus in familial Parkinson disease. Am J Hum Genet 2008; 82 : 822–33. [Google Scholar]
- Helmer C, Pasquier F, Dartigues JF. Épidémiologie de la maladie d’Alzheimer et des syndromes apparentés. Med Sci (Paris) 2006; 22 : 288–96. [Google Scholar]
- Checler F, Alves da Costa C, Dumanchin-Njock C, et al. Métabolisme du précurseur du peptide amyloïde et présénilines. Med Sci (Paris) 2002; 18 : 717–22. [Google Scholar]
© 2009 médecine/sciences - Inserm / SRMS
Liste des tableaux
Maladies associées à des mutations pathogènes des gènes impliqués dans la cascade de signalisation de l’insuline et de l’IGF-1. (http://www.orphanet.com). a : récepteur de l’insuline ; b : récepteur de l’IGF-1 ; c : growth factor receptor-bound protein 10 ; d : Grb-10 interacting GYF protein 2 ; e : hypoglycémie à jeun ; f : retard de croissance intra-utérin ; ND : non déterminé lors de nos travaux détaillés dans [10].
Liste des figures
|
Figure 1. Les voies de signalisation de l’insuline et de l’IGF-1 et des facteurs de croissance neuronale. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.