")
")
| Issue |
Med Sci (Paris)
Volume 22, Number 6-7, Juin-Juillet 2006
|
|
|---|---|---|
| Page(s) | 573 - 575 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20062267573 | |
| Published online | 15 juin 2006 | |
Hypocrétines : entre l’envie et le besoin… ou l’émergence d’une nouvelle voie d’origine hypothalamique impliquée dans la motivation et l’addiction
Hypocretins: between desire and needs… Toward the understanding of a new hypothalamic brain pathway involved in motivation and addiction
Centre de Neurosciences Psychiatriques-Service Universitaire de l’Enfant et de l’Adolescent, Hôpital de Cery, CH-1008 Prilly-Lausanne, Suisse
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
L’hypothalamus est l’interprète des besoins de l’organisme (garant de l’homéostasie du milieu interne) et l’architecte des adaptations périphériques. Siège du dialogue neuroendocrinien, il contribue à la régulation de la faim, de la soif, de la température corporelle, en plus de son rôle de modulation de l’alternance veille/sommeil. Plus spécifiquement, c’est l’hypothalamus latéral (LH) qui joue un rôle crucial dans la régulation homéostasique des besoins vitaux de l’organisme. Le LH est en effet considéré comme un maillon essentiel dans la régulation des comportements motivés ; sa lésion entraîne une nette diminution de la recherche et de la consommation d’eau et de nourriture chez le rat. En outre, Olds et Milner [1] ont mis en évidence que des rats étaient capables de fournir un effort considérable pour auto-stimuler électriquement leur LH. La démonstration, également reproduite chez l’homme [2], fait concevoir le LH comme une des pièces maîtresses de la fonction de récompense cérébrale (FRC)1, fonction qui conditionne les aspects motivationnels de toute stratégie comportementale et pousse à la réalisation d’un objectif donné. Toutefois, les mécanismes moléculaires impliqués dans la régulation de la fonction de récompense hypothalamique demeurent largement méconnus.
Historiquement, un neuromédiateur a été associé à la FRC et à la motivation : la dopamine (DA) [3]. De nombreux auteurs ont confirmé l’étroite corrélation entre libération de dopamine et comportements motivés (recherche d’eau, de nourriture, de partenaire sexuel). On considère aujourd’hui que la dopamine confère une valeur particulière à un stimulus reconnu comme pertinent par rapport à l’ensemble des sollicitations sensorielles, émotionnelles ou affectives, ce que l’on résume par la notion de gain signal/bruit [3].
Drogues et besoins naturels
Comme les drogues et les besoins naturels mettent en jeu la même circuiterie cérébrale2, une implication du système DA a été clairement établie dans le phénomène addictif [3] sans pour autant apporter de réponses thérapeutiques novatrices au cours des 30 dernières années.
L’addiction (pour une drogue) est un trouble compulsif chronique qui se caractérise par une envie irrépressible de rechercher et de consommer une substance en quantité importante, malgré les conséquences délétères sur la santé et l’intégration sociale. Le taux de récidive très élevé après une période d’abstinence/désintoxication marque le douloureux constat des limites de nos connaissances et de nos moyens thérapeutiques. Ainsi, l’identification de structures cérébrales responsables de la vulnérabilité au risque de rechute est cruciale pour le développement de nouveaux traitements susceptibles d’enrayer le cycle infernal de l’addiction.
Hypocrétines : hypervigilance, stress et équilibre énergétique
L’hypocrétine-1 et -2 (Hcrt-1 et Hcrt-2) [4] (ou orexine-A et -B [5]) sont deux neuropeptides synthétisés par quelques milliers de neurones localisés dans le LH. Les neurones à Hcrt projettent toutefois dans l’ensemble du cerveau, notamment en direction des structures cérébrales impliquées dans la modulation de la motivation, de la récompense, du stress et de l’éveil (Figure 1). Les premières observations concernant ce système lui ont attribué un rôle dans la régulation de l’appétit, sans que cette hypothèse fasse l’unanimité, alors que chacun reconnaît aujourd’hui le lien étroit entre dégénérescence du système Hcrt et narcolepsie [6]. Ainsi, l’hypothèse actuelle confère au système Hcrt un rôle pivot entre intégration des besoins caloriques et orchestration du niveau de vigilance [7]. Conscients du profil incomparable du système Hcrt, tant par son anatomie que par sa participation dans la régulation de la fonction corticotrope et de l’ensemble des systèmes de neuromédiateurs (dopamine, noradrénaline, sérotonine, acétylcholine) [7], nous avons les premiers suggéré un rôle de l’Hcrt dans la modulation de la FRC [8].
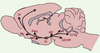 |
Figure 1. Coupe sagittale d’un cerveau de rat schématisant l’organisation du système hypocrétinergique (Hcrt). Les corps cellulaires Hcrt sont localisés exclusivement dans l’hypothalamus latéral (LH) mais les neurones Hcrt projettent dans l’ensemble du cerveau. CP : cortex préfrontal ; NAc : noyau accumbens ; Amyg : amygdale ; BNST : noyau du lit de la strie terminale ; ATV : aire tegmentaire ventrale ; LC/LTg : locus cœruleus/ tegmentum latéral ; NR : noyau du raphé ; LDT/PPT : tegmentum latérodorsal/tegmentum pédonculopontin. |
Quand on cherche la dopamine, on la trouve…
Trois équipes ont récemment proposé une implication du système Hcrt dans l’addiction via la sollicitation du système DA. Brièvement, il a été montré une étroite corrélation entre activation des neurones Hcrt du LH dans le cerveau de rats et manifestation d’une préférence marquée pour le compartiment d’une boîte précédemment associée avec une récompense (nourriture ou drogue) [9]. L’activation pharmacologique des neurones Hcrt précipite un comportement de recherche chez des rats qui ne manifestaient plus cette préférence [9]. La même rechute se manifeste après injection d’Hcrt-1 directement dans l’aire tegmentaire ventrale (ATV) [9, 10]. En complément, il a été établi que l’Hcrt-1 jouait un rôle critique dans la plasticité synaptique des neurones glutamatergiques de l’ATV après traitement chronique à la cocaïne, et que l’administration d’un antagoniste des récepteurs Hcrt bloquait le phénomène de sensibilisation comportementale à la cocaïne [11]. Enfin, les souris mutantes déficientes en Hcrt ne manifestent aucune préférence dans un test de préférence conditionnée avec de la morphine [10].
…mais la dopamine n’explique pas tout
La notion de sensibilisation comportementale aux effets (principalement locomoteurs) de la cocaïne continue de faire couler beaucoup d’encre. Si le rôle de la DA dans ce phénomène s’avère particulièrement bien étayé, la pertinence de cette observation pour expliquer, et a fortiori comprendre, le trouble addictif dans son ensemble est d’avantage sujet à caution. Première interrogation, les observations neuroanatomiques ne permettent pas de reconnaître au système Hcrt une capacité d’innervation importante de l’ATV, du moins par comparaison à l’innervation du locus cœruleus, du noyau du lit de la strie terminale ou du noyau du raphé [7]. Deuxièmement, il est parfaitement établi que le système DA n’est pas un pré-requis nécessaire à la manifestation d’un comportement de préférence dans un paradigme de préférence conditionnée [12]. En revanche, les neuromédiateurs impliqués dans la réponse au stress - notamment les systèmes à noradrénaline (NA) et à corticolibérine (CRH) - ont un rôle parfaitement établi dans les phénomènes de résurgence d’un comportement de recherche de drogue après une période d’abstinence.
Telle a été notre approche pour l’étude de l’implication du système Hcrt dans les mécanismes neurobiologiques responsables de la vulnérabilité au risque de rechute [13]. Contrairement aux autres équipes, nous avons étudié le comportement de rats libres de s’auto-administrer de la cocaïne. Nous avons montré que l’Hcrt ne modulait ni la prise, ni la motivation pour prendre de la cocaïne. Toutefois, nous avons établi qu’une injection intracérébroventriculaire (ICV) d’Hcrt entraînait de façon dose-dépendante la résurgence d’un comportement d’appuis frénétiques sur le levier préalablement associé avec la délivrance de drogue après une période d’abstinence3 (Figure 2). En bloquant au préalable les systèmes NA et CRH, nous sommes parvenus à empêcher une telle rechute. Enfin, contrairement aux traitement pharmacologiques entraînant une libération de DA (cocaïne, morphine…), l’administration d’Hcrt entraîne une élévation des seuils d’autostimulation chez le rat. Observation cruciale car l’abaissement du seuil d’autostimulation reflète la dynamisation de la FRC, ce qui sous-tend, ou du moins contribue, à l’état d’excitation/euphorie associé à la prise de drogue. Dans le cas présent, la détérioration du fonctionnement de la FRC induit par l’Hcrt s’accorde davantage avec le seuil d’exigence de satisfaction, très élevé, mesuré au cours d’une période de sevrage, d’un stress ou après administration de CRH. II exclut de fait toute participation directe de la DA. Afin de compléter notre démonstration, nous avons pu prouver qu’un traitement avec un antagoniste des récepteurs Hcrt bloquait complètement toute récidive dans un modèle de rechute induit par un état de stress.
 |
Figure 2. Évolution temporelle du nombre d’appuis sur le levier associé à la délivrance de cocaïne. Au cours de la phase d’auto-administration, chaque appui sur le levier est récompensé par une injection intraveineuse de cocaïne. Ensuite, au cours de la phase d’extinction, chaque appui sur le levier n’est plus renforcé par la délivrance de cocaïne. Enfin, survient la résurgence du comportement d’appui sur le levier induit par l’administration d’hypocrétine (Hcrt), bien que la cocaïne ne soit plus délivrée. |
Conclusions
En résumé, deux visions s’opposent sur les mécanismes neurobiologiques responsables des effets induits par l’administration d’Hcrt chez des animaux « intoxiqués » à la cocaïne ou à la morphine. Qu’il s’agisse d’un effet appétitif sous-tendu par une stimulation du système DA ou qu’il s’agisse d’une réaction de stress qui pousse l’animal à trouver une solution pour pallier le désagrément qu’il subit, il est clair que le système Hcrt joue un rôle de premier plan dans la symptomatologie liée au phénomène addictif. Pour notre part, nous estimons que ce système est à l’interface des réseaux de neurones impliqués dans la régulation de l’hypervigilance, du stress et de la motivation. En tant que tel, il nous paraît concevable de corréler activation hypocrétinergique et excitation mentale. En ce sens, nous considérons que ce système pourrait être impliqué dans le phénomène de craving (envie irrépressible de se procurer et de consommer une substance), et qu’une altération de ce système pourrait représenter un facteur de vulnérabilité au risque de rechute chez des toxicomanes abstinents et les alcooliques [14]. De fait, cibler le système Hcrt représente une innovation thérapeutique théoriquement susceptible de diminuer le risque de récidive, facteur emblématique de toute politique de prévention en matière d’addiction.
Erratum
Dans l’article de Pierre H. Vachon, « Survie cellulaire : différences et différenciation » (m/s n° 4, vol. 22, avril 2006, pages 423-9) :
page 425, dans la Figure 2, il fallait d’une part lire Bcl-2 et non Bcl, et RSK-1 et non RSK, d’autre part.
page 428, lignes 4 à 6, il fallait lire : …les entérocytes indifférenciés ont une moindre susceptibilité face à l’anoïkose que les entérocytes différenciés, et non pas …les entérocytes différenciés ont une moindre susceptibilité face à l’anoïkose que les entérocytes indifférenciés.
Remerciements
L’auteur tient à remercier ses proches collaborateurs sans lesquels il n’aurait pu accomplir ce travail : Paul J. Kenny, Athina Markou, George F. Koob et Luis de Lecea (The Scripps Research Institute, Californie, USA). Il remercie également les Professeurs Pierre J. Magistretti (Centre de Neurosciences Psychiatriques) et Olivier Halfon (Service Universitaire de l’Enfant et de l’Adolescent, Lausanne, Suisse) pour leurs encouragements amicaux et leur soutien financier, ainsi que la Fondation pour la Recherche Médicale.
La FRC peut grossièrement se comparer au corrélat neurobiologique de la recherche de satisfaction et/ou de gratification, c’est-à-dire la volonté d’obtenir/d’assouvir un objet de satisfaction, état d’esprit souvent lié à une certaine forme d’exaltation, voire d’excitation.
On considère que les drogues piratent le fonctionnement normal du cerveau et détournent le système de récompense cérébrale dans leur propre intérêt, de sorte que l’idéation de la drogue représente l’unique source d’intérêt et de motivation, le reste n’ayant plus d’importance.
On parle plus spécifiquement d’une période d’extinction du comportement d’appui sur le levier précédemment associé à la drogue.
Références
- Olds J, Milner P. Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain. J Comp Physiol Psychol 1954; 47 : 419–27. [Google Scholar]
- Bishop MP, Elder ST, Heath RG. Intracranial self-stimulation in man. Science 1963; 140 : 394–6. [Google Scholar]
- Wise RA. Dopamine, learning and motivation. Nat Rev Neurosci 2004; 5 : 483–94. [Google Scholar]
- De Lecea L, Kilduff TS, Peyron C, et al. The hypocretins : hypothalamus-specific peptides with neuroexcitatory activity. Proc Natl Acad Sci USA 1998; 95 : 322–7. [Google Scholar]
- Sakurai T, Amemiya A, Ishii M, et al. Orexins and orexin receptors : a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998; 92 : 573–85. [Google Scholar]
- Boutrel B, Koob GF. What keeps us awake : the neuropharmacology of stimulants and wakefulness-promoting medications. Sleep 2004; 27 : 1181–94. [Google Scholar]
- Sutcliffe JG, De Lecea L. The hypocretins : setting the arousal threshold. Nat Rev Neurosci 2002; 3 : 339–49. [Google Scholar]
- Boutrel B, Kenny PJ, Markou A, Koob GF. Hypocretin and the brain reward function. In : De Lecea L, Sutcliffe JG, eds. The hypocretins : integrators of physiological signals. New York : Springer, 2005:345–55. [Google Scholar]
- Harris GC, Wimmer M, Jones GA. A role for lateral hypothalamic orexin neurons in reward seeking. Nature 2005; 437 : 556–9. [Google Scholar]
- Narita M, Nagumo Y, Hashimoto S, et al. Direct involvement of orexinergic systems in the activation of the mesolimbic dopamine pathway and related behaviors induced by morphine. J Neurosci 2006; 26 : 398–405. [Google Scholar]
- Borgland SL, Taha SA, Sarti F, et al. Orexin A in the VTA is critical for the induction of synaptic plasticity and behavioral sensitization to cocaine. Neuron 2006; 49 : 589–601. [Google Scholar]
- Hnasko TS, Sotak BN, Palmiter RD. Morphine reward in dopamine-deficient mice. Nature 2005; 438 : 854–57. [Google Scholar]
- Boutrel B, Kenny PJ, Specio SE, et al. Role for hypocretin in mediating stress-induced reinstatement of cocaine-seeking behavior. Proc Natl Acad Sci USA 2005; 102 : 19168–73. [Google Scholar]
- Lawrence AJ, Cowen MS, Yang HJ, et al. The orexin system regulates alcohol-seeking in rats. Br J Pharmacol 2006; 5 juin online. [Google Scholar]
© 2006 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Coupe sagittale d’un cerveau de rat schématisant l’organisation du système hypocrétinergique (Hcrt). Les corps cellulaires Hcrt sont localisés exclusivement dans l’hypothalamus latéral (LH) mais les neurones Hcrt projettent dans l’ensemble du cerveau. CP : cortex préfrontal ; NAc : noyau accumbens ; Amyg : amygdale ; BNST : noyau du lit de la strie terminale ; ATV : aire tegmentaire ventrale ; LC/LTg : locus cœruleus/ tegmentum latéral ; NR : noyau du raphé ; LDT/PPT : tegmentum latérodorsal/tegmentum pédonculopontin. |
| Dans le texte | |
|
Figure 2. Évolution temporelle du nombre d’appuis sur le levier associé à la délivrance de cocaïne. Au cours de la phase d’auto-administration, chaque appui sur le levier est récompensé par une injection intraveineuse de cocaïne. Ensuite, au cours de la phase d’extinction, chaque appui sur le levier n’est plus renforcé par la délivrance de cocaïne. Enfin, survient la résurgence du comportement d’appui sur le levier induit par l’administration d’hypocrétine (Hcrt), bien que la cocaïne ne soit plus délivrée. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.