")
")
| Issue |
Med Sci (Paris)
Volume 22, Number 4, Avril 2006
|
|
|---|---|---|
| Page(s) | 361 - 363 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2006224361 | |
| Published online | 15 avril 2006 | |
La méthylation des histones n’est plus ce qu’elle était
Histone demethylation unravelled ?
Institut de Recherche Interdisciplinaire, Cnrs FRE 2963, IRI @ Institut de Biologie de Lille, 1, rue du Professeur Calmette, 59021 Lille Cedex, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Dans le noyau, l’ADN eucaryote s’associe avec les histones (H2A, H2B, H3 et H4) pour former l’unité fondamentale de la chromatine : le nucléosome. L’architecture précise de la chromatine détermine si celle-ci est permissive ou non à la transcription et à d’autres événements qui dépendent de son organisation, tels que la réplication, la réparation de l’ADN ou la recombinaison. Alors que la grande majorité des nucléosomes sont constitués des mêmes histones, une immense diversité de nucléosomes différents découle des modifications post-traductionnelles qui touchent les histones. Celles-ci peuvent être acétylées, méthylées, phosphorylées, ubiquitinylées ou sumoylées. Il est proposé que ces différentes modifications agissent en combinaison pour former un « code histone » [1–3]. Ce code est lu par des protéines non-histones qui se lient aux résidus modifiés des histones, et influencent alors l’organisation de la chromatine, la transcription ou la réplication. Certaines modifications comme l’acétylation ou la phosphorylation sont réversibles et dynamiques, et souvent associées à la régulation inductible de gènes individuels. En revanche, pendant de nombreuses années, la méthylation des histones était considérée comme une marque épigénétique stable et irréversible qui rendait compte du caractère héritable des états ouvert (euchromatine) ou fermé (hétérochromatine) de la chromatine. Cette vision découlait de l’incapacité à mettre en évidence une activité histone déméthylase et des études qui suggéraient que la demi-vie de la méthylation des histones est similaire à celle des histones elles-mêmes [4].
Cependant, le dogme de l’irréversibilité de la méthylation des histones est maintenant remis en question. Des études ont montré que la méthylation pouvait même être extrêmement dynamique. Par exemple, la méthylation de la lysine 9 de l’histone H3 des nucléosomes localisés au niveau de gènes qui répondent à l’inflammation disparaît lors de leur activation, puis est restaurée lors de la répression transcriptionnelle post-induction [5]. Depuis fin 2004, un nouveau pas a été franchi avec la découverte des protéines et des mécanismes directement impliqués dans la déméthylation des histones (Figure 1).
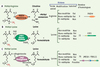 |
Figure 1. Mécanismes enzymatiques de la déméthylation et spécificité de substrat des histone déméthylases. A. La méthylation des arginines est supprimée par les arginine désiminases (PADI) pour engendrer une citrulline et du méthylammonium. Les enzymes PADI sont peu spécifiques et sont actives sur différentes arginines monométhylées des histones. Elles peuvent même convertir des arginines non-méthylées en citrullines. B. Deux mécanismes rendent compte de la déméthylation des méthyl-lysines. Les amine oxydases (de type LSD1) régénèrent une lysine et du formaldéhyde. La formation d’une imine intermédiaire par transfert de deux atomes d’hydrogène sur le cofacteur FAD nécessite un azote protoné, ce qui limite la réaction aux lysines mono- et diméthylées. C. Les di-oxygénases à domaine JmjC engendrent également une lysine et du formaldéhyde mais peuvent théoriquement déméthyler les tri-méthyl-lysines. À la différence des histone déméthylases spécifiques des arginines, les histones déméthylases spécifiques des lysines ont une très grande spécificité de substrat : LSD1 pour les lysines 4 ou 9 de l’histone H3, selon le cofacteur associé, et FBLX11 pour la lysine 36 de l’histone H3. Le nombre de protéines humaines qui possèdent des domaines désiminases, amine oxidases ou JmjC similaires à ceux de PADI4, LSD1 ou FBXL11 est indiqué (http://www.sanger.ac.uk/Software/Pfam/). Seule la déméthylation des monométhyl-lysine est montrée (adapté de [14]). |
Les agents de déméthylation des histones
Les arginines sont sujettes à une conversion enzymatique qui engendre un acide aminé non conventionnel. La peptidylarginine désiminase 4 humaine (PADI4/PAD4) convertit les arginines monométhylées des histones H3 et H4 en citrulline par déméthylimination [6, 7]. Le fait que PADI4 transforme également les arginines non méthylées en citrullines, qui ne peuvent plus être méthylées, suggère que le rôle de PADI4 serait d’éliminer les histones en tant que substrats des arginine méthyltransférases, et non seulement de déméthyler les histones.
LSD1 (lysine-specific demethylase 1) est une amine oxydase nucléaire dépendante de l’adénine flavine dinucléotide (FAD) capable de déméthyler la lysine 4 de l’histone H3 [8]. LSD1 catalyse une réaction oxydative de l’amine via le clivage oxydatif de la liaison α-CH du substrat pour former une imine en réduisant un cofacteur, la flavine. L’imine intermédiaire est ensuite hydrolysée spontanément pour produire du carbinolamine, groupe instable qui se transforme en formaldéhyde, régénérant ainsi une lysine déméthylée. Seules les lysines mono- et di-méthylées sont des substrats de LSD1. En effet, la formation de l’imine intermédiaire nécessite une lysine protonée, et les lysines tri-méthylées ne peuvent donc pas être substrats des amine oxydases. L’activité et la spécificité de déméthylation des lysines 4 de l’histone H3 dépend à la fois des autres modifications post-traductionnelles de l’histone H3, comme son état de phosphorylation en sérine 10 [9] et de la présence de cofacteurs. En effet, quand LSD1 est associé au cofacteur CoREST, LSD1 déméthyle la lysine 4 de l’histone H3 [10] (une marque de l’euchromatine), alors qu’associée avec le récepteur des androgènes, LSD1 déméthyle la lysine 9 de l’histone H3 [11] (une marque de l’hétérochromatine). Ainsi, la protéine LSD1 agit à la fois comme un co-activateur ou un co-répresseur transcriptionnel.
Plus récemment, il a été proposé [12, 13] que la déméthylation des lysines pouvait se faire par hydroxylation des groupes méthyles. Cette réaction est catalysée par le domaine JmjC qui possède une activité di-oxygénase dépendante de l’α-cétoglutarate et du fer divalent. La réaction utilise le Fe(II) pour activer une molécule de di-oxygène et former une molécule hautement réactive d’oxoferryl [Fe(IV)O], qui peut hydroxyler la méthyl-lysine ; la déméthylation produit alors du formaldéhyde. Les premières histone déméthylases de ce type caractérisées sont JHMD1A (FBXL11) et JHMD1B (FBXL10) qui déméthylent spécifiquement la lysine 36 de l’histone H3 sous sa forme mono- ou di-méthylée, mais pas sous sa forme tri-méthylée [13].
Alors que la découverte des premières histone déméthylases représente une avancée majeure dans la biologie de la chromatine et de l’épigénétique, elle soulève également de nombreuses questions. La méthylation des arginines peut être le résultat d’une induction hormonale, mais si leur déméthylation est associée à l’apparition d’une citrulline, comment une nouvelle induction peut-elle avoir lieu ? La citrulline constitue-t-elle une nouvelle marque du code histone ? Les amine oxidases (environ 20 membres) et les protéines à domaine JmjC (environ 70 membres) répondent en termes de nombre à la complexité de la méthylation des lysines contrôlée par plus de 70 protéines à motif méthyltransférase spécifique (le domaine SET). Toutefois, aucune protéine capable de déméthyler les tri-méthyl-lysines n’a encore été identifiée, bien que la réaction d’hydroxylation des groupes méthyles en soit théoriquement capable. Et si la méthylation des histones est aussi dynamique, quels sont les mécanismes responsables de l’héritabilité de l’organisation de la chromatine et de son maintien au cours des mitoses successives ?
Références
- Strahl BD, Allis CD. The language of covalent histone modifications. Nature 2000; 403 : 41–5. [Google Scholar]
- Jenuwein T, Allis CD. Translating the histone code. Science 2001; 293 : 1074–80. [Google Scholar]
- Ray-Gallet D, Gérard A, Polo S, Almouzni G. Variations sur le thème du « code histone ». Med Sci (Paris) 2005; 21 : 384–9. [Google Scholar]
- Byvoet P, Shepherd GR, Hardin JM, Noland BJ. The distribution and turnover of labelled methyl groups in histone fractions of cultured mammalian cells. Arch Biochem Biophys 1972; 148 : 558–67. [Google Scholar]
- Saccani S, Natoli G. Dynamic changes in histone H3 Lys 9 methylation occurring at tightly regulated inducible inflammatory genes. Genes Dev 2002; 16 : 2219–24. [Google Scholar]
- Cuthbert GL, Daujat S, Snowden AW, et al. Histone deimination antagonizes arginine methylation. Cell 2004; 118 : 545–53. [Google Scholar]
- Wang Y, Wysocka J, Sayegh J, et al. Human PAD4 regulates histone arginine methylation levels via demethylimination. Science 2004; 306 : 279–83. [Google Scholar]
- Shi Y, Lan F, Matson C, et al. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 2004; 119 : 941–53. [Google Scholar]
- Forneris F, Binda C, Vanoni MA, et al. Human histone demethylase LSD1 reads the histone code. J Biol Chem 2005; 280 : 41360–5. [Google Scholar]
- Shi Y, Matson C, Lan F, et al. Regulation of LSD1 histone demethylase activity by its associated factors. Mol Cell 2005; 19 : 857–64. [Google Scholar]
- Metzger E, Wissmann M, Yin N, et al. LSD1 demethylates repressive histone marks to promote androgen-receptor-dependent transcription. Nature 2005; 437 : 436–9. [Google Scholar]
- Trewick SC, McLaughlin PJ, Allshire RC. Methylation : lost in hydroxylation ? EMBO Rep 2005; 6 : 315–20. [Google Scholar]
- Tsukada Y, Fang J, Erdjument-Bromage H, et al. Histone demethylation by a family of JmjC domain-containing proteins. Nature 2006; 439 : 811–6. [Google Scholar]
- Kubicek S, Jenuwein T. A crack in histone lysine methylation. Cell 2004; 119 : 903–6. [Google Scholar]
© 2006 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Mécanismes enzymatiques de la déméthylation et spécificité de substrat des histone déméthylases. A. La méthylation des arginines est supprimée par les arginine désiminases (PADI) pour engendrer une citrulline et du méthylammonium. Les enzymes PADI sont peu spécifiques et sont actives sur différentes arginines monométhylées des histones. Elles peuvent même convertir des arginines non-méthylées en citrullines. B. Deux mécanismes rendent compte de la déméthylation des méthyl-lysines. Les amine oxydases (de type LSD1) régénèrent une lysine et du formaldéhyde. La formation d’une imine intermédiaire par transfert de deux atomes d’hydrogène sur le cofacteur FAD nécessite un azote protoné, ce qui limite la réaction aux lysines mono- et diméthylées. C. Les di-oxygénases à domaine JmjC engendrent également une lysine et du formaldéhyde mais peuvent théoriquement déméthyler les tri-méthyl-lysines. À la différence des histone déméthylases spécifiques des arginines, les histones déméthylases spécifiques des lysines ont une très grande spécificité de substrat : LSD1 pour les lysines 4 ou 9 de l’histone H3, selon le cofacteur associé, et FBLX11 pour la lysine 36 de l’histone H3. Le nombre de protéines humaines qui possèdent des domaines désiminases, amine oxidases ou JmjC similaires à ceux de PADI4, LSD1 ou FBXL11 est indiqué (http://www.sanger.ac.uk/Software/Pfam/). Seule la déméthylation des monométhyl-lysine est montrée (adapté de [14]). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.