")
")
| Issue |
Med Sci (Paris)
Volume 21, Number 5, Mai 2005
|
|
|---|---|---|
| Page(s) | 455 - 457 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2005215455 | |
| Published online | 15 May 2005 | |
Qu’apprend-on du génome de Legionella pneumophila?
What do we learn from the genome of Legionella pneumophila?
Unité de Génomique des micro-organismes pathogènes, CNRS URA 2171, Institut Pasteur, 28, rue du Docteur Roux, 75724 Paris Cedex 15, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Legionella pneumophila, l’agent responsable de la légionellose (Figure 1), doit son nom à une épidémie survenue en 1976 chez 182 participants du 58e congrès de la Légion américaine à Philadelphie. Depuis, de nombreuses épidémies de légionellose ont été décrites en Amérique du Nord et en Europe : on estime entre 8 000 et 18 000 le nombre de cas de légionellose chaque année aux États-Unis. En France, alors que des cas sporadiques sont régulièrement recensés, huit épidémies particulièrement graves ont été identifiées depuis 1998. À ce jour, la légionellose reste donc une maladie grave, avec une incidence annuelle réelle estimée en France à un minimum de 1 200 cas avec une mortalité de 20 à 30 %.
 |
Figure 1. Legionella pneumophila. Son flagelle permet à la bactérie d’être mobile (microscopie électronique). |
Les légionelles sont des bactéries vivant en milieu aquatique et affectionnant particulièrement les eaux tièdes. Elles parasitent habituellement les amibes, protozoaires qui prolifèrent dans l’eau, mais sont également capables d’infecter l’homme par le biais d’aérosols. Les bactéries peuvent atteindre les alvéoles pulmonaires où elles sont phagocytées par les macrophages. Contrairement à d’autres bactéries qui sont tuées par ces macrophages, les légionelles sont capables de s’y multiplier et de les lyser, causant une pneumopathie grave.
Malgré des avancées récentes sur la compréhension de la réplication et la virulence de Legionella, ce pathogène humain restait l’un des derniers dont la séquence génomique n’était pas encore déterminée. Afin d’élucider et caractériser les bases génétiques de la virulence de L. pneumophila, nous avons adopté une approche génomique en déterminant la séquence complète du génome de deux souches de L. pneumophila, responsables d’épidémies de légionellose en France [1]. Une troisième souche a été séquencée aux États-Unis [2]. Ces trois souches appartiennent au sérogroupe 1 qui est responsable de plus de 85 % des cas. La souche dite « Paris » est la seule souche endémique connue à ce jour : alors que les autres souches provoquent des cas dans une région limitée, la souche « Paris » est associée à des cas de légionellose partout en France et peut-être même en Europe [3]. Elle est ainsi responsable de 12,7 % des cas de légionellose en France et de 33 % des cas dans la région parisienne ; elle était notamment à l’origine de l’épidémie de l’hôpital Georges Pompidou. La deuxième souche étudiée dite « Lens » avait causé, entre novembre 2003 et janvier 2004, l’épidémie la plus importante connue en France avec 86 cas et 17 morts. La troisième souche est dérivée de la souche identifiée à Philadelphie lors de l’épidémie de 1976.
L’analyse et la comparaison des génomes mettent en évidence une grande diversité et plasticité de l’espèce L. pneumophila et fournissent des bases pour comprendre la pathogénicité de Legionella, en particulier pour déchiffrer les interactions hôte-pathogène.
Caractéristiques générales des trois génomes
Le génome de L. pneumophila consiste en un chromosome circulaire d’environ 3,7 Mb avec une composition de l’ADN en bases G-C de 38 %. Chacune des souches possède également un ou plusieurs plasmides. L’analyse des séquences prédit la présence d’environ 3 000 gènes dont 80 % codent pour des protéines possédant des homologues (orthologues) dans d’autres organismes. Certains de ces orthologues ont des fonctions connues (60 % des gènes), ce qui permet de prédire cette même fonction dans L. pneumophila.
Exploitation des fonctions de la cellule hôte
Le génome de Legionella reflète la co-évolution de cette bactérie avec des cellules eucaryotes comme les protozoaires dans l’eau et les macrophages pulmonaires lors de l’infection humaine. En effet, nous avons identifié 62 gènes codant pour des protéines ayant une similarité avec des protéines eucaryotes, plus qu’il n’en a jamais été décrit dans les génomes bactériens déjà séquencés. Beaucoup possèdent notamment des motifs d’interaction protéiques retrouvés essentiellement chez des eucaryotes. La présence de ces protéines suggère que Legionella est capable d’interagir avec son hôte et de moduler les fonctions de la cellule hôte pour les détourner à son avantage (Figure 2).
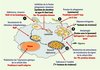 |
Figure 2. Cycle intracellulaire de Legionella dans le macrophage et protéines qui pourraient interférer à des différents stades. En bleu, sont indiquées les étapes connues et importantes dans le cycle infectieux de L. pneumophila. En rouge, sont indiquées les nouvelles protéines identifiées qui pourraient interférer dans ces étapes du cycle cellulaire. |
Les protéines EnhC et LidL contiennent chacune un motif eucaryote Sel1 et interviennent respectivement dans l’invasion et le trafic intracellulaire [4, 5]. Trois autres protéines, avec des domaines Sel1, ont été identifiées par le séquençage et pourraient également intervenir dans des interactions hôte-pathogène.
Pour survivre à l’ingestion par les macrophages, Legionella inhibe la fusion phagosome-lysosome et échappe ainsi à la dégradation par le lysosome. Puis elle survit et se réplique, protégée dans une vacuole dérivée du réticulum endoplasmique. La protéine RalF, qui contient un motif eucaryote Sec7, intervient à ce stade du cycle cellulaire en recrutant le réticulum endoplasmique à la surface de la vacuole. Par ailleurs, trois protéines de Legionella sont similaires aux sérine/ thréonine protéine kinases eucaryotes ; chez les procaryotes, ces protéines peuvent jouer des rôles divers tels que l’inhibition de la fusion phagosome-lysosome ou la modulation des défenses de l’hôte [6].
Un autre exemple de protéines qui pourraient interférer avec des fonctions de l’hôte sont trois protéines à motif F-box que nous avons identifiées dans le génome de Legionella. Cette famille de protéines est principalement impliquée dans l’ubiquitinylation et la lyse consécutive de protéines promotrices ou inhibitrices du cycle cellulaire par dégradation protéasomique [7]. Donc ces protéines de Legionella sont probablement capables d’interagir avec la machinerie d’ubiquitinylation de la cellule hôte. Jusqu’à présent, on n’avait encore identifié qu’une seule protéine à motif F-box chez un procaryote (Agrobacterium tumefaciens).
Finalement, la présence d’autres gènes codant pour des protéines typiquement eucaryotes : protéines à motifs U-box, sphingosine-1-phosphate lyase, sphingosine kinase and apyrase suggère que Legionella peut moduler directement le cycle cellulaire de l’hôte.
Plasticité du génome de Legionella
Chaque souche possède 9 à 13 % de protéines spécifiques, pourcentage important pour trois souches d’une même espèce. Elles possèdent, entre autres, un ou plusieurs plasmides spécifiques qui pourraient intervenir dans la virulence et la plasticité de Legionella. Des études ont déjà montré une corrélation entre la présence de plasmides et la virulence [8] ainsi qu’une plus grande persistance dans l’environnement des souches possédant des plasmides [9]. Un plasmide portant un système de sécrétion de type IV impliqué dans la virulence, qui peut être présent à la fois sous forme excisée ou intégrée dans le chromosome, a été identifié dans les souches L. pneumophila « Paris » et « Philadelphia ». Dans d’autres souches, le même type d’élément instable est associé à la virulence : les souches perdent la virulence lorsque le plasmide est en position excisée [10].
Conclusions
L’analyse du génome de Legionella a montré la grande diversité de l’espèce L. pneumophila et a permis de prédire l’implication d’un grand nombre de protéines dans des interactions hôte-pathogène. Des analyses fonctionnelles de ces gènes confirmeraient si ces protéines sont utilisées par L. pneumophila pour détourner les fonctions de la cellule hôte, amibe ou macrophage pulmonaire humain, à son avantage. La connaissance du génome devrait également permettre, à long terme, de mettre en place de meilleurs outils de diagnostic, de nouvelles armes thérapeutiques ainsi que des biocides plus actifs que ceux qui sont connus à ce jour pour la décontamination de l’eau, et plus particulièrement des systèmes d’eau chaude.
Références
- Cazalet C, Rusniok C, Brüggeman H, et al. Evidence in the Legionella pneumophila genome for exploitation of host cell functions and high genome plasticity. Nat Genet 2004; 36 : 1165–73. [Google Scholar]
- Chien M, Morozova I, Shi S, et al. The genomic sequence of the accidental pathogen Legionella pneumophila. Science 2004; 305 : 1966–8. [Google Scholar]
- Aurell H, Farge P, Meugnier H, et al. Legionella pneumophila serogroup 1 strain Paris : endemic distribution throughout France. J Clin Microbiol 2003; 41 : 3320–2. [Google Scholar]
- Cirillo SLG, Lum J, Cirillo JD. Identification of a novel loci involved in entry by Legionella pneumophila. Microbiology 2000; 146 : 1345–59. [Google Scholar]
- Conover GM, Derre I, Vogel JP, Isberg RR. The Legionella pneumophila LidA protein : a translocated substrate of the Dot/Icm system associated with the maintenance of bacterial integrity. Mol Microbiol 2003; 48 : 305–21. [Google Scholar]
- Walburger A, Koul A, Ferrari G, et al. Protein kinase G from pathogenic mycobacteria promotes survival within macrophages. Science 2004; 304 : 1800–4 [Google Scholar]
- Craig KL, Tyers M. The F-box : a new motif fot ubiquitin dependent proteolysis in cell cycle regulation and signal transduction. Prog Biophys Mol Biol 1999; 72 : 299–328. [Google Scholar]
- Bezanson G, Fernandez R, Haldane D, et al. Virulence of patient and water isolates of Legionella pneumophila in guinea pigs and mouse L929 cells varies with bacterial genotype. Can J Microbiol 1994; 40 : 426–31. [Google Scholar]
- Brown A, Vickers RM, Elder EM, et al. Plasmid and surface antigen markers of endemic and epidemic Legionella pneumophila strains. J Clin Microbiol 1982; 16 : 230–35. [Google Scholar]
- Luneberg E, Mayer B, Daryab N, et al. Chromosomal insertion and excision of a 30 kb unstable genetic element is responsible for phase variation of lipopolysaccharide and other virulence determinants in Legionella pneumophila. Mol Microbiol 2001; 39 : 1259–71. [Google Scholar]
© 2005 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Legionella pneumophila. Son flagelle permet à la bactérie d’être mobile (microscopie électronique). |
| Dans le texte | |
|
Figure 2. Cycle intracellulaire de Legionella dans le macrophage et protéines qui pourraient interférer à des différents stades. En bleu, sont indiquées les étapes connues et importantes dans le cycle infectieux de L. pneumophila. En rouge, sont indiquées les nouvelles protéines identifiées qui pourraient interférer dans ces étapes du cycle cellulaire. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.