")
")
| Issue |
Med Sci (Paris)
Volume 20, Number 1, Janvier 2004
|
|
|---|---|---|
| Page(s) | 7 - 8 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20042017 | |
| Published online | 15 janvier 2004 | |
Les protéines se liant à l’ADN méthylé : interprètes du code épigénétique
DNA methyltransferases: from chromatin remodeling to cancer
Cnrs UMR 218, Institut Curie, Section de Recherche, 26, rue d’Ulm, 75248 Paris Cedex 05, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
L’hérédité repose sur la transmission fidèle de séquences d’acides nucléiques d’un organisme à ses descendants. L’information résidant dans la séquence peut toutefois être étendue par des modifications chimiques du matériel génétique, stables et transmissibles d’une génération à la suivante. Chez les eucaryotes, la méthylation des cytosines en est un exemple particulièrement bien documenté. Le groupement méthyle a un caractère épigénétique car il est transmis aux molécules filles après la réplication par des enzymes conservant cette méthylation. La Figure 1 montre la position de groupements méthyle dans le grand sillon de l’ADN au niveau d’un dinucléotide CpG.
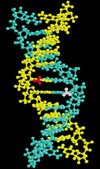 |
Figure 1. Structure de l’ADN au niveau d’un dinucléotide CpG méthylé. Les groupements méthyle des deux chaînes (en violet et en rouge) se situent dans le grand sillon, en position accessible. Après la réplication de l’ADN, les sites hémiméthylés sont complétés par une méthyltransférase de maintien, ce qui assure la transmission de ce motif. |
La méthylation des cytosines n’existe pas chez tous les organismes. Saccharomyces cerevisiae et Caenorhabditis elegans en sont par exemple totalement dépourvus. Chez les espèces où elle est présente, la méthylation des cytosines est souvent associée à la formation de l’hétérochromatine (→), l’inhibition de la recombinaison homologue, et la répression de l’expression des gènes [1]. D’autres fonctions liées à la méthylation de l’ADN sont spécifiques d’un groupe d’organismes. C’est ainsi le cas de l’inactivation du chromosome X, qui n’a lieu que chez les mammifères.
(→) m/s 2003, n° 12, p. 1181
Dans le règne animal les cytosines méthylées se trouvent presque invariablement dans le dinucléotide CpG. Chez les vertébrés, ces sites méthylables suivent une distribution non uniforme : il existe des domaines, appelés îlots CpG, où ce dinucléotide est plus fortement représenté. Les îlots CpG correspondent fréquemment au promoteur et au premier exon des gènes et ils ne sont en général pas méthylés dans les cellules saines. En revanche dans les cellules cancéreuses, il est fréquent d’observer des méthylations aberrantes des îlots CpG, corrélées à la répression transcriptionnelle des gènes qui leur sont associés. Même si l’origine du phénomène est débattue, l’importance de la méthylation dans la progression des cancers est bien établie (→) ; il y a donc un enjeu important dans la compréhension de la répression des gènes imposée par la méthylation de l’ADN.
(→) m/s 2003, n° 4, p. 477
La méthylation exerce une action directe sur l’expression des gènes en empêchant la fixation de protéines qui contrôlent la transcription. Ainsi les activateurs transcriptionnels Sp1, CREB, YY1, et d’autres encore, ne se lient plus à certaines de leurs cibles si elles sont méthylées. Mais les conséquences de la méthylation peuvent aussi être indirectes et nécessiter l’intervention d’interprètes. A ce titre, il existe des protéines capables de lier spécifiquement l’ADN méthylé et de traduire ce signal en terme d’expression génique.
Chez les animaux, une famille de protéines reconnaît les CpG méthylés par un motif conservé de 70 acides aminés appelé MBD (methyl-CpG binding domain). Chez les vertébrés, cette famille se compose de cinq membres : MeCP2, MBD1, MBD2, MBD3 et MBD4. MeCP2 est le membre fondateur de la famille, les autres ayant été identifiés en 1998 par la recherche d’homologies de séquence avec cette protéine dans les bases de données [2].
MeCP2, MBD1, MBD2 et MBD3 sont associées à des complexes répresseurs de la transcription. Les mécanismes de la répression ne sont pas parfaitement compris mais ils impliquent notamment des modifications locales de la chromatine [3]. Parmi ces protéines, MDB3 est un membre à part car elle ne peut se lier intrinsèquement aux CpG méthylés, sa spécificité et son mode d’action sur la régulation des gènes restent donc à préciser.
L’ajout du groupement méthyle sur les cytosines augmente la probabilité de mutation. En effet, la cytosine méthylée peut subir une désamination spontanée qui la transforme en thymine, créant ainsi un mésappariement T/G dans l’ADN. La protéine MBD4 est capable de reconnaître ce mésappariement et d’exciser la thymine par une activité glycosylase. D’autres systèmes ont ensuite pour tâche de réparer le site abasique. Bien que MBD4 soit capable de reconnaître les CpG méthylés, son affinité pour les mésappariements T/G est plus forte, et elle semble exercer surtout un rôle de surveillance dans les génomes fortement méthylés.
Récemment la famille MBD s’est agrandie grâce à l’identification de six nouveaux membres par analyse de séquence [4]. On ne sait pas encore si ces protéines sont capables de lier les CpG méthylés, mais elles offrent de nouvelles perspectives de régulation des gènes. Ces interprètes potentiels de la méthylation seront vraisemblablement les derniers mis en évidence dans la famille MBD chez les mammifères. Les séquences complètes des génomes de l’homme et de la souris permettent en effet aux recherches d’être exhaustives.
En dehors de la famille des protéines MBD, une seule autre protéine capable de reconnaître l’ADN méthylé a été découverte chez les vertébrés [5]. Cette protéine, appelée Kaiso, reconnaît spécifiquement les CpG méthylés par un domaine de liaison à l’ADN contenant trois doigts de zinc. Le motif en doigt de zinc est très courant chez tous les organismes et permet généralement des interactions spécifiques avec l’ADN. La possibilité théorique qu’une protéine à doigt de zinc puisse lier les cytosines méthylées avait été remarquée en 1998 [6]. Cependant, en l’absence d’exemples naturels, la pertinence biologique de cette observation n’était pas claire. Le génome humain renferme deux gènes dont le domaine à doigts de zinc est très proche de celui de Kaiso, qui pourraient donc potentiellement lier l’ADN méthylé. Les études structurales de Kaiso et de ses homologues promettent ainsi de définir les propriétés d’un nouveau domaine de liaison à l’ADN méthylé.
De façon inattendue, une étude récente montre que Kaiso peut aussi reconnaître une séquence spécifique non méthylée, de sept nucléotides, par l’intermédiaire du même domaine à doigts de zinc [7]. Puisque le domaine à doigts de zinc de Kaiso reconnaît une séquence d’ADN ainsi que les CpG méthylés il n’y a pas incompatibilité entre ces deux spécificités. Il est donc permis d’imaginer que, parmi les protéines à doigts de zinc dont le site consensus de liaison a été défini, se cachent d’autres interprètes de la méthylation. Plus généralement, il est possible que des protéines se lient à l’ADN méthylé par d’autres motifs de liaison à l’ADN, par exemple le motif hélice-tour-hélice. Enfin, puisque la méthylation est présente dans une vaste gamme d’organismes et qu’elle est associée à des fonctions diverses, il est envisageable que des protéines sans homologues chez les vertébrés reconnaissent spécifiquement l’ADN méthylé.
Il semble raisonnable d’espérer que l’identification de ces interprètes secrets de la méthylation offrira de nombreuses potentialités en biotechnologie et en thérapeutique, et apportera aussi sa contribution dans les domaines de l’évolution et de la biologie fondamentale.
Références
- Colot V, Rossignol JL. Eukaryotic DNA methylation as an evolutionary device. Bioessays 1999; 21 : 402–11. [Google Scholar]
- Hendrich B, Bird A. Identification and characterization of a family of mammalian methyl-CpG binding proteins. Mol Cell Biol 1998; 18 : 6538–47. [Google Scholar]
- Fuks F. DNA methyltransferases : from chromatin remodeling to cancer. Med Sci 2003; 19 : 477–80. [Google Scholar]
- Roloff TC, Ropers HH, Nuber UA. Comparative study of methyl-CpG-binding domain proteins. BMC Genomics 2003; 4 : 1. [Google Scholar]
- Prokhortchouk A, Hendrich B, Jorgensen H, et al. The p120 catenin partner Kaiso is a DNA methylation-dependent transcriptional repressor. Genes Dev 2001; 15 : 1613–8. [Google Scholar]
- Choo Y. Recognition of DNA methylation by zinc fingers. Nat Struct Biol 1998; 5 : 264–5. [Google Scholar]
- Daniel JM, Spring CM, Crawford HC, Reynolds AB, Baig A. The p120(ctn)-binding partner Kaiso is a bi-modal DNA-binding protein that recognizes both a sequence-specific consensus and methylated CpG dinucleotides. Nucleic Acids Res 2002; 30 : 2911–9. [Google Scholar]
© 2004 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Structure de l’ADN au niveau d’un dinucléotide CpG méthylé. Les groupements méthyle des deux chaînes (en violet et en rouge) se situent dans le grand sillon, en position accessible. Après la réplication de l’ADN, les sites hémiméthylés sont complétés par une méthyltransférase de maintien, ce qui assure la transmission de ce motif. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.