")
")
| Issue |
Med Sci (Paris)
Volume 19, Number 11, Novembre 2003
|
|
|---|---|---|
| Page(s) | 1054 - 1056 | |
| Section | Le Magazine : Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/200319111054 | |
| Published online | 15 novembre 2003 | |
Le fuseau mitotique est-il un organe sensoriel de la cellule ?
Mitotic spindle dictates the symmetry of division
Institut de Biochimie, École polytechnique fédérale de Zurich, ETH-Zentrum Universitatstrasse 16,8092 Zurich, Suisse
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
La génétique a établi un paradoxe que nous ne finissons pas de résoudre: alors que toutes les cellules de l’organisme sont génétiquement identiques, elles adoptent des fonctions et des morphologies distinctes. Les progrès de la biologie cellulaire et de la biologie du développement suggèrent qu’un même processus, la mitose, assure à la fois la prolifération et la diversification cellulaires.
Au cours de la mitose, la cellule assemble un fuseau mitotique et un appareil de clivage pour ensuite se diviser. Le fuseau a deux fonctions essentielles:il sépare en deux lots identiques les chromatides, qui ont subi préalablement un processus de réplication, et il positionne l’appareil de clivage entre ces deux lots de chromosomes.
Bien que les deux cellules filles soient génétiquement identiques, elles peuvent être structurellement différentes [1]. Cela est dû au fait que les cellules eucaryotes sont souvent polarisées, certains de leurs composants étant distribués préférentiellement à l’un ou l’autre pôle de la cellule (→) Lorsqu’une cellule ainsi polarisée se divise perpendiculairement à son axe de polarité (Figure 1A, A”), les composants cellulaires ne se distribuent pas de manière équivalente dans les deux cellules filles, qui développent alors des propriétés différentes:on parle de division asymétrique. Si, en revanche, la cellule se divise parallèlement à son axe de polarité (Figure 1A, A’), les deux cellules filles qui en sont issues seront identiques. Puisque c’est le fuseau qui dirige le positionnement de l’appareil de clivage, c’est son orientation qui détermine si la division cellulaire est symétrique ou asymétrique. Cependant, nous savons encore très peu de choses sur le mécanisme d’orientation du fuseau mitotique dans la cellule.
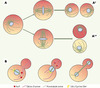 |
Figure 1. Positionnement du fuseau mitotique et son rôle dans les divisions asymétriques. A. Schéma représentatif des cellules animales. La cellule représentée est polarisée, ce qu’indique le gradient de couleur. Selon l’axe de division de la cellule, qui est déterminé par l’axe d’orientation du fuseau, deux cellules identiques (A’) ou deux cellules distinctes (A’’) sont produites. B. Orientation du fuseau dans la cellule de levure S. cerevisiae. La protéine Kar9 se localise à l’extrémité des microtubules émanant d’un seul des deux pôles du fuseau. Cette protéine permet la migration des extrémités des microtubules le long des câbles d’actine. Comme ces cables proviennent tous du cortex du bourgeon, cela aboutit à l’orientation du fuseau le long de l’axe mèrebourgeon. Cette orientation assure une bonne ségrégation des chromosomes entre la cellule mère et la cellule fille pendant l’anaphase. |
(→) m/s2003, n°6-7, p.656
La mitose est l’un des processus les plus anciens dans l’histoire de la vie - sans mitose il n’y aurait aucune cellule - et n’est donc pas une propriété propre aux organismes multicellulaires. Et c’est effectivement l’étude d’un organisme unicellulaire, la levure de bière, qui nous apporte le plus d’informations sur les mécanismes d’orientation du fuseau. Cet organisme prolifère par bourgeonnement et les mitoses qu’il effectue sont systématiquement asymétriques. Pendant la formation du bourgeon, qui émerge de la surface de la cellule mère, le fuseau mitotique s’aligne le long de l’axe mère-bourgeon et répartit les chromosomes entre ces deux compartiments (Figure 1B). Lorsqu’il a reçu son lot de chromosomes, le bourgeon se sépare de la mère, donnant naissance à deux cellules distinctes, une grosse mère et une fille, petite. De nombreux mutants affectant la capacité de la cellule à positionner correctement son fuseau mitotique ont été isolés et l’étude des gènes impliqués est très informative.
Le fuseau est constitué à ses pôles de structures appelées spindle pole bodies(SPB, équivalents des centrosomes des cellules animales), et il est formé de différents types de microtubules. Les microtubules astraux, qui émergent des SPB en direction de la périphérie cellulaire, sont directement impliqués dans le positionnement du fuseau de la levure. La protéine Kar9, qui est fonctionnellement apparentée à la protéine APC (adenomatous polyposis coli) impliquée dans le cancer du côlon chez l’humain, est aussi un facteur clé pour l’orientation du fuseau [2]. Kar9 s’associe indirectement aux microtubules via la protéine EB1 [3]. Compte tenu de ces affinités moléculaires, nous avons montré que Kar9 s’accumulait de façon préférentielle à l’extrémité des microtubules [4, 5]. De façon remarquable, Kar9 ne s’accumulait pas à l’extrémité de tous les microtubules, mais seulement de ceux qui émanent du «vieux» SPB [4]. Ce SPB est celui qui s’oriente en direction du bourgeon pendant la métaphase et entre dans le bourgeon lors de l’anaphase. En l’absence de Kar9, aucun des deux pôles du fuseau n’est dirigé vers le bourgeon et le fuseau ne s’aligne pas le long de l’axe mère-bourgeon. Ces résultats indiquent que Kar9 n’est recruté que sur certains microtubules, et qu’il guide vers le bourgeon le SPB auquel ces microtubules sont attachés. Comment expliquer que Kar9 ne se lie pas à tous les microtubules, et que Kar9 ne dirige qu’un seul SPB vers le bourgeon ?
L’étude des modifications de Kar9 nous a permis de répondre à la première question [4]. En effet, Kar9 est une protéine phosphorylée dans la cellule, et cette phosphorylation interfère avec sa capacité à lier EB1, et donc à s’associer aux microtubules. La kinase qui phosphoryle Kar9 n’est autre que la kinase dépendante des cyclines, Cdk1, qui dirige l’assemblage du fuseau. La forme de Cdk1 qui phosphoryle Kar9 est localisée sur le jeune SPB. Les mutations qui, d’une façon ou d’une autre, empêchent la phosphorylation de Kar9 ont pour conséquence la localisation de Kar9 au bout des microtubules émanant des deux SPB. Ces données nous permettent de nous faire une idée assez claire du mécanisme par lequel la localisation de Kar9 est restreinte aux microtubules émanant spécifiquement du vieux SPB: l’asymétrie de Kar9 est due au fait que le processus de recrutement de Kar9 sur les microtubules naissants est inhibé par Cdk1 sur le « jeune »SPB, mais pas sur le « vieux » [4]. Mais la raison pour laquelle Cdk1 s’accumule préférentiellement sur un SPB, et pas sur l’autre, n’est pas encore bien claire.
La réponse à la seconde question - comment Kar9 dirige-t-elle le «vieux» SPB vers le bourgeon ? - nous a été apportée par deux observations. L’une, biochimique, indique que Kar9 a la capacité d’interagir avec une myosine, la myosine de type V, et de la recruter sur les microtubules [6, 7]. Les myosines transforment l’énergie chimique de l’ATP en activité motrice le long de câbles d’actine. La seconde observation est que les microtubules émanant du vieux SPB, et seulement ceux-ci, s’orientent dans la cellule pour aller contacter le cortex du bourgeon [4]. Cette orientation dépend de la présence de câbles d’actine émanant du cortex du bourgeon, de la présence de Kar9 et de l’interaction de Kar9 avec la myosine V. En conséquence, les formes non phosphorylables de Kar9, qui sont chargées sur les microtubules émanant des deux pôles du fuseau, dirigent les deux SPB vers le bourgeon. Le fuseau n’est alors plus orienté parallèlement à l’axe de polarité de la cellule, mais perpendiculairement. Cette orientation dépend de l’activité de la myosine. Donc, en distribuant Kar9 de façon symétrique ou asymétrique, la cellule peut décider de diviser son noyau parallèlement ou perpendiculairement à son axe de symétrie.
En résumé, un modèle émerge de ces données, selon lequel le fuseau agirait comme une sorte de boussole intracellulaire. Tout d’abord, comme l’aiguille de la boussole, le fuseau est asymétrique. L’asymétrie de la boussole est magnétique, celle du fuseau est chimique. Dans le cas de la boussole, l’asymétrie de l’aiguille lui permet de s’orienter dans un champ magnétique. Pour le fuseau, son asymétrie lui permet de s’orienter grâce à une molécule motrice dans un champ de câbles d’actine, si ces câbles sont eux-mêmes orientés. Le fuseau semble donc fonctionner comme une sorte d’organe sensoriel, qui est capable de percevoir l’orientation de la cellule et de transmettre cette information à l’appareil de division.
Références
- Kaltschmidt JA, Brand AH. Asymmetric cell division:microtubule dynamics and spindle asymmetry. J Cell Sci 2002; 115: 2257–64. [Google Scholar]
- Miller RK, Rose MD. Kar9p is a novel cortical protein required for cytoplasmic microtubule orientation in yeast. J Cell Biol 1998;140: 377–90. [Google Scholar]
- Miller RK, Cheng SC, Rose MD. Bim1p/Yeb1p mediates the Kar9p-dependent cortical attachment of cytoplasmic microtubules. Mol Biol Cell 2000; 11: 2949–59. [Google Scholar]
- Liakopoulos D, Kusch J, Grava S, Vogel J, Barral Y. Asymmetric loading of Kar9 onto spindle poles and microtubules ensures proper spindle alignment. Cell 2003; 112: 561–74. [Google Scholar]
- Kusch J, Meyer A, Snyder MP, Barral Y. Microtubule capture by the cleavage apparatus is required for proper spindle positioning in yeast. Genes Dev 2002; 16: 1627–39. [Google Scholar]
- Yin H, Pruyne D, Huffaker TC, Bretscher A. Myosin V orientates the mitotic spindle in yeast. Nature 2000; 406: 1013–5. [Google Scholar]
- Hwang E, Kusch J, Barral Y, Huffaker TC. Spindle orientation in Saccharomyces cerevisiae depends on the transport of microtubule ends along polarized actin cables. J Cell Biol 2003; 161: 483–8. [Google Scholar]
© 2003 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Positionnement du fuseau mitotique et son rôle dans les divisions asymétriques. A. Schéma représentatif des cellules animales. La cellule représentée est polarisée, ce qu’indique le gradient de couleur. Selon l’axe de division de la cellule, qui est déterminé par l’axe d’orientation du fuseau, deux cellules identiques (A’) ou deux cellules distinctes (A’’) sont produites. B. Orientation du fuseau dans la cellule de levure S. cerevisiae. La protéine Kar9 se localise à l’extrémité des microtubules émanant d’un seul des deux pôles du fuseau. Cette protéine permet la migration des extrémités des microtubules le long des câbles d’actine. Comme ces cables proviennent tous du cortex du bourgeon, cela aboutit à l’orientation du fuseau le long de l’axe mèrebourgeon. Cette orientation assure une bonne ségrégation des chromosomes entre la cellule mère et la cellule fille pendant l’anaphase. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.