")
")
Figure 3.
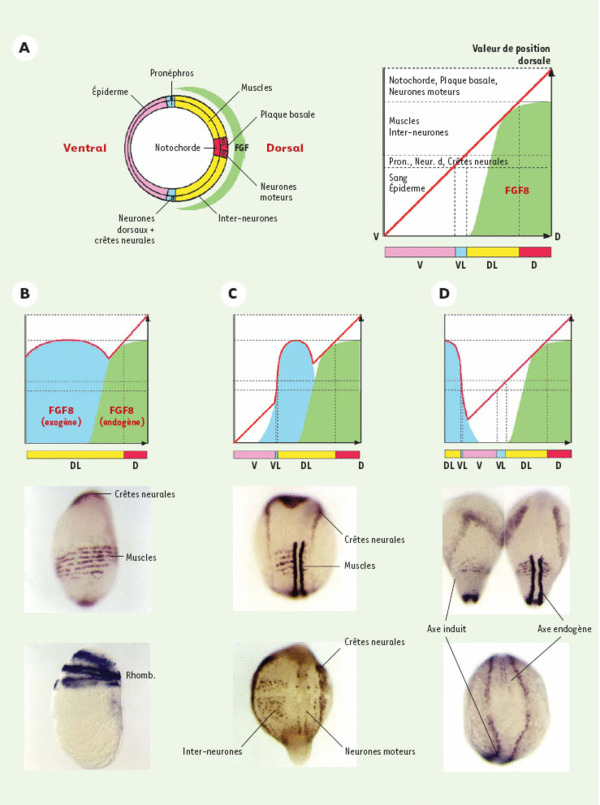
Manipulations de l’axe D/V via le contrôle de l’activité morphogénique des BMP. A. Section d’un embryon suivant l’axe D/V au stade gastrula permettant de mettre en évidence l’organisation des tissus mésodermiques et ectodermiques le long de cet axe. On peut ainsi distinguer les territoires dorsaux (D, en rouge) comprenant la notochorde dans le feuillet mésodermique, la plaque basale du tube neural et les neurones moteurs dans l’ectoderme ; les territoires dorsaux latéraux (DL, en jaune) comprenant les muscles pour le mésoderme, les inter-neurones pour l’ectoderme ; les territoires ventraux latéraux (VL, en bleu) comprenant le pronéphros pour le mésoderme ; les neurones dorsaux et les crêtes neurales pour l’ectoderme ; les territoires ventraux (V, en rose) comprenant le sang pour le mésoderme et l’épiderme pour le feuillet ectodermique. Le croissant vert représente le gradient d’expression du FGF8 au stade gastrula. On peut définir les différents territoires le long de l’axe D/V en fonction d’une valeur de position dorsale. Selon ce modèle, les différents territoires sont formés en réponse à la valeur de position dorsale que mesure chaque cellule le long de l’axe D/V. L’information positionnelle dorsale, nulle sur la face ventrale de l’embryon et maximale sur la face dorsale (courbe rouge) représente la résultante de l’interaction entre les BMP ventralement et leurs inhibiteurs dorsaux. En fonction de la valeur de cette information de position, les cellules vont adopter un sort ventral (rose), ventro-latéral (bleu), dorso-latéral (jaune) ou dorsal (rouge). La pertinence de ce modèle est illustrée par l’étude de l’inhibition de l’expression des BMP par le FGF8 dans des expériences de surexpression. La surface verte sur ce graphe correspond à la contribution du FGF8 à l’établissement de l’axe D/V (fondé sur son territoire d’expression dorsal et dorso-latéral, ainsi que sur son activité biologique : capable d’induire des structures dorso-latérales mais incapables d’induire les sorts les plus dorsaux). B. Lorsque l’embryon est saturé en FGF8 (injection d’ARN sens au stade une cellule), l’expression des BMP est réprimée dans tout l’embryon et en conséquence toutes les cellules adoptent un sort dorso-latéral caractérisé par la circularisation du territoire musculaire (embryon du haut) ou des rhombomères (Rhomb) du cerveau postérieur (embryon du bas). Ce phénotype est identique au phénotype de perte de fonction des BMP présenté en figure 2B. En pareil cas, la courbe de l’information positionnelle (rouge) est déplacée vers les valeurs correspondant aux territoires dorso-latéraux pour toutes les cellules à l’exception de celles des territoires dorsaux endogènes. C. Lorsque le clone sécrétant le FGF8 est en position latérale on observe un élargissement des territoires dorsolatéraux tant dans le mésoderme (élargissement unilatéral des sillons somitiques) que dans l’ectoderme (élargissement unilatéral de la plaque neurale bordée par les cellules des crêtes neurales ; augmentation unilatérale du nombre d’inter-neurones). D. Lorsque le clone est en position ventrale, il conduit à la formation d’un axe secondaire partiellement défectif comprenant les territoires dorsolatéraux mais pas les territoires dorsaux (notochorde et partie ventrale du tube neural). L’embryon du haut est photographié sous ses deux faces, dorsale (axe endogène) et ventrale (axe induit). Les mêmes résultats peuvent être observés en inhibant l’activité des BMP par la surexpression d’autres facteurs inhibiteurs qui tels Noggin et Chordin agissent en fixant les BMP les empêchant ainsi de stimuler leur récepteur spécifique.
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.