")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 436 - 439 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026076 | |
| Published online | 29 mai 2026 | |
La glutamine au cœur du métabolisme énergétique des macrophages dans l’athérosclérose
Glutamine at the heart of macrophage energy metabolism in atherosclerosis
Inserm U1065, Université Côte d’Azur, Centre méditerranéen de médecine moléculaire (C3M), Institut hospitalo-universitaire RespirERA, Nice, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
L’athérosclérose est la cause majeure des maladies cardiovasculaires, qui sont responsables de plus de 50 % des décès dans les pays industrialisés. Elle se caractérise par la formation de plaques d’athérome riches en lipides dans la paroi artérielle, associée à une inflammation chronique. Cette inflammation lésionnelle résulte notamment d’un dysfonctionnement des macrophages, qui empêche la résolution de l’inflammation et la réparation vasculaire. L’exposition prolongée à une surcharge alimentaire lipidique et glucidique, typique des modes de vie industrialisés, instaure un environnement inflammatoire favorisant la progression des plaques d’athérome [1, 2]. Il n’est donc pas surprenant que les hyperlipidémies et l’hyperglycémie constituent des facteurs de risque cardio-métaboliques majeurs, ciblés par les stratégies thérapeutiques actuelles. À ce contexte métabolique défavorable s’ajoute souvent un déséquilibre protéique, conséquence d’une alimentation ultra-transformée et d’un apport journalier supérieur à la recommandation (0,8 g/kg/jour). La glutamine (Gln), acide aminé le plus abondant dans le plasma, joue un rôle clé dans l’homéostasie métabolique et immunitaire, notamment chez les personnes âgées [1], et fait l’objet de recherches récentes quant à son implication dans l’athérosclérose.
La glutamine : interface métabolique entre la « nutrigénétique » et l’athérosclérose
La concentration plasmatique de glutamine (Gln) est maintenue par un équilibre dynamique entre les apports alimentaires, son stockage et sa resynthèse dans le muscle squelettique, et son utilisation hépatique par désamination, assurant ainsi un recyclage métabolique continu entre la Gln et l’acide glutamique (Glu). Dans une étude épidémiologique fondée sur le suivi prospectif de 862 individus de la cohorte multiethnique MESA (multiethnic study of atherosclerosis), le rapport Gln/Glu plasmatique était positivement corrélé à la progression de l’athérosclérose [3], avec une ampleur comparable à celle de la glycémie. Un rapport Gln/Glu supérieur à 5 a ainsi été proposé comme nouveau facteur de risque de l’athérosclérose [3]. Des analyses pangénomiques d’association (genome-wide association study, GWAS) couplées à des analyses métabolomiques ont permis d’identifier des déterminants génétiques de la concentration plasmatique de Gln dans le gène codant la glutaminase 2 (GLS2). Cette enzyme est majoritairement produite dans le foie, où elle catalyse la désamination de Gln. Pour tenter de mettre en évidence une relation causale entre ces variants génétiques et les maladies cardiovasculaires, une approche de « randomisation mendélienne »1 a été utilisée dans un groupe de personnes atteintes d’athérosclérose des artères coronaires, en utilisant comme variable instrumentale un polymorphisme génétique de GLS2 (rs2657879) influant sur l’activité de l’enzyme [3]. Cette analyse a confirmé l’existence d’un lien causal entre ce polymorphisme de GLS2, la concentration plasmatique de Gln, et l’athérosclérose, un lien qui a pu être validé dans des modèles d’athérosclérose chez la souris, dans lesquels l’invalidation de GLS2 dans le foie augmente le rapport Gln/Glu plasmatique ainsi que la progression des plaques d’athérosclérose associée à un remodelage matriciel et inflammatoire [3]. Pour différencier la part génétique de la part alimentaire, une supplémentation en Gln a également été réalisée dans des modèles murins déficients ou non en GLS2. Ces études nutrigénétiques ont permis de mettre en évidence l’importance de la désamination hépatique de Gln pour compenser un excès d’apport alimentaire et prévenir le développement de l’athérosclérose [3] (Figure 1).
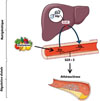 |
Figure 1 Nutrigénétique de la glutamine et dialogue entre foie et vaisseaux dans l’athérosclérose. La concentration plasmatique de la glutamine (Gln) résulte d’un équilibre entre l’apport alimentaire de Gln, qui augmente sa disponibilité, et sa désamination hépatique par la glutaminase 2 (GLS2), qui convertit Gln en acide glutamique (Glu). Des polymorphismes (single nucleotide polymorphism, SNP) du gène codant GLS2 modulent ce flux métabolique et le rapport Gln/Glu (Gln/Glu ratio, GGR) dans le sang. Ils définissent un nouvel axe nutrigénétique pour contrer une surconsommation de Gln et limiter son accumulation systémique. Un GGR supérieur à 5 est proposé comme un nouveau facteur de risque cardiovasculaire associé à un remodelage matriciel et une inflammation à distance des plaques d’athérosclérose. |
La plasticité des flux de glutamine : signature inflammatoire de l’athérosclérose
La glutamine est considérée comme un carburant majeur du système immunitaire, en soutenant les processus anaplérotiques2 mitochondriaux indispensables à la production d’énergie sous forme d’ATP. Cependant, différents flux cellulaires de Gln induisent des phénotypes macrophagiques distincts [4, 5]. Le métabolisme des macrophages activés par des lipopolysaccharides bactériens repose sur la glycolyse aérobie et sur la conversion de Gln en acide succinique via un shunt métabolique « GABAergique » pour soutenir la réponse pro-inflammatoire. Au contraire, les macrophages réparateurs, induits par l’interleukine-4, augmentent leur absorption de Gln, qui est dirigée vers un shunt métabolique différent du précédent, appelé « transamination non canonique », afin de maintenir l’oxydation phosphorylante et la production d’ATP et ainsi soutenir la réponse anti-inflammatoire réparatrice via l’efférocytose [4]. Dans des conditions réparatrices, l’augmentation de l’absorption de Gln par les macrophages est due à l’induction de l’expression du transporteur SLC7A7 qui favorise son entrée dans la cellule. L’inhibition expérimentale de ce transporteur réduit la respiration mitochondriale, la production d’ATP, l’expression de marqueurs réparateurs tels que CD206, ainsi que l’efférocytose, mimant le phénotype observé lors d’une privation de Gln ou en l’absence de la glutaminase 1 (GLS1), l’enzyme limitante catalysant la conversion de Gln en Glu dans les macrophages [5]. Ainsi, l’absorption de la Gln via SLC7A7 agit comme un « rhéostat métabolique » dont le rôle est essentiel pour l’homéostasie énergétique et les fonctions réparatrices des macrophages. Cette flexibilité métabolique, indispensable à l’adaptation fonctionnelle des macrophages, est fortement altérée dans le contexte de l’athérosclérose. En effet, bien qu’on constate une augmentation de l’expression de SLC7A7 dans les plaques d’athérosclérose carotidiennes instables, elle s’accompagne d’une forte diminution de l’expression de GLS1, suggérant une redirection du flux de Gln avec une perte de glutaminolyse. Cette perturbation métabolique pourrait expliquer la corrélation inverse entre l’expression de GLS1 et la taille des plaques ainsi que leur composition en corps nécrotiques, car la disponibilité de Gln pour soutenir la production mitochondriale d’ATP serait compromise, limitant ainsi l’élimination des lipides [4, 5]. Ainsi dans des modèles murins d’athérosclérose, la restriction soit de l’influx de Gln via SLC7A7, soit de la glutaminolyse via GLS1, entraîne une accumulation de macrophages inflammatoires et enrichis en lipides (macrophages spumeux TREM2+), associée à une perte d’efférocytose dans les lésions. Ces altérations favorisent la progression des lésions vers un phénotype plus instable, caractérisé par une accumulation de lipides, une augmentation des débris cellulaires et la formation d’un cœur nécrotique [4, 5]. En exploitant le modèle murin d’hyperglutaminémie induite par la déficience hépatique en GLS2, nous avons montré que l’incorporation de Gln par les macrophages est dirigée vers des voies métaboliques alternatives, notamment la biosynthèse des hexosamines, au détriment de la production de Glu, de la respiration mitochondriale et de la production d’ATP [5]. Cette reprogrammation s’accompagne d’une activation transcriptionnelle marquée par des signatures inflammatoires et de remodelage de la matrice e xtracellulaire, conduisant à une altération de la chape fibreuse de la plaque athéromateuse [5]. Ainsi, la Gln ne constitue pas uniquement un carburant métabolique, mais agit également comme un véritable centre de signalisation orchestrant les réponses métaboliques et fonctionnelles des macrophages au cours de l’athérosclérose (Figure 2).
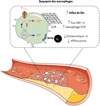 |
Figure 2 Fluctuation des flux de glutamine et dyspepsie des macrophages dans l’athérosclérose. L’augmentation du flux de glutamine (Gln), illustrée par l’imagerie TEPglutamine, est associée à une augmentation de l’expression du transporteur SLC7A7 par les macrophages de la plaque d’athérosclérose. Cependant, du fait de la diminution de l’expression de la glutaminase 1 (GLS1) dans ces macrophages, les processus anaplérotiques mitochondriaux indispensables à la production d’ATP sont diminués, conduisant à une perte de remodelage des membranes et à un défaut d’efférocytose (« dyspepsie macrophagique »). Le flux de Gln alimente alors préférentiellement la voie de biosynthèse des hexosamines (hexosamine biosynthesis pathway, HBP), ce qui conduit à un remodelage de la matrice extracellulaire (extracellular matrix, ECM). Ces fluctuations métaboliques de la glutamine au sein des macrophages de la plaque d’athérosclérose entraînent sa progression. |
La TEP-glutamine : un marqueur métabolique des macrophages athéromateux
Des fluctuations répétées de la glycémie ou de la lipidémie, liées aux habitudes alimentaires, altèrent le fonctionnement immunitaire et contribuent aux complications vasculaires associées à la méta-inflammation [6, 7]. Cette dépendance métabolique explique l’utilisation d’analogues radioactifs du glucose pour détecter l’activité inflammatoire des plaques d’athérome par tomographie par émission de positons (TEP-scan). Toutefois, l’efficacité de cette technique est limitée par la captation simultanée du glucose par la moelle osseuse, site de production des cellules de la réaction inflammatoire [8, 9], et par la consommation basale de glucose par les cardiomyocytes, à l’origine d’un « bruit de fond ». Ces contraintes soulignent la nécessité d’identifier des traceurs métaboliques plus spécifiques. Des travaux récents utilisant des analogues de Gln marqués par 18F chez l’Homme ont révélé une incorporation beaucoup plus sélective dans les macrophages des plaques d’athérome comparativement au glucose [5, 10]. Cette captation est corrélée avec l’expression du transporteur SLC7A7 et l’accumulation de macrophages spumeux TREM2+ [10], conformément aux constats précédents dans les modèles murins [5]. Bien que cette imagerie du flux de Gln constitue un biomarqueur prometteur de l’inflammation des plaques athéromateuses, il est nécessaire d’approfondir notre compréhension des flux métaboliques cellulaires sous-jacents afin d’ouvrir la voie à des applications thérapeutiques.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Yvan-Charvet L, Barouillet T, Borowczyk C. Haematometabolism rewiring in atherosclerotic cardiovascular disease. Nat Rev Cardiol 2025 ; 6 : 414-30. [Google Scholar]
- Janssen H, Koekkoek LL, Swirski FK. Effects of lifestyle factors on leukocytes in cardiovascular health and disease. Nat Rev Cardiol 2024 ; 3 : 157-69. [Google Scholar]
- Murcy F, Borowczyk C, Gourion-Arsiquaud S, et al. GLS2 links glutamine metabolism and atherosclerosis by remodeling artery walls. Nat Cardiovasc Res 2024 ; 12 : 1454-67. [Google Scholar]
- Merlin J, Ivanov S, Dumont A, et al. Non-canonical glutamine transamination sustains efferocytosis by coupling redox buffering to oxidative phosphorylation. Nat Metab 2021 ; 3 : 1313-26. [Google Scholar]
- Benhmammouch S, Borowczyk C, Pierrot-Blanchet C, et al. Slc7a7 licenses macrophage glutaminolysis for restorative functions in atherosclerosis. Nat Metab 2025 ; 7 : 1924-38. [Google Scholar]
- Flynn MC, Kraakman MJ, Tikellis C, et al. Transient intermittent hyperglycemia accelerates atherosclerosis by promoting myelopoiesis. Circ Res 2020 ; 127 : 877-92. [Google Scholar]
- Takaoka M, Zhao X, Lim HY, et al. Early intermittent hyperlipidaemia alters tissue macrophages to fuel atherosclerosis. Nature 2024 ; 634 : 457-65. [Google Scholar]
- Sarrazy V, Viaud M, Westerterp M, et al. Disruption of Glut1 in hematopoietic stem cells prevents myelopoiesis and enhanced glucose flux in atheromatous plaques of ApoE-/- mice. Circ Res 2016 ; 118 : 1062-77. [Google Scholar]
- Morioka S, Perry JSA, Raymond MH, et al. Efferocytosis induces a novel SLC program to promote glucose uptake and lactate release. Nature 2018 ; 563 : 714-8. [Google Scholar]
- Örd T, Palani S, Giroud Gerbetant J, et al. Single-cell to pre-clinical evaluation of Trem2, Folr2, and Slc7a7 as macrophage-associated biomarkers for atherosclerosis. Cardiovasc Res 2025 ; 121 : 2503-19. [Google Scholar]
Dans ce type d’analyse, les variants génétiques associés à un facteur de risque pressenti (ici, la concentration plasmatique de Gln comme facteur de risque de l’athérosclérose), parce qu’ils sont attribués de façon aléatoire au zygote dès sa conception, sont utilisés à la place de ce facteur de risque pour limiter l’influence de possibles « facteurs de confusion » empêchant l’interprétation causale d’une différence statistiquement significative entre les groupes dans une étude « non randomisée ».
Cet adjectif qualifie une réaction chimique qui produit un métabolite, c’est-à-dire une espèce chimique intermédiaire d’une voie métabolique (source : Wikipédia).
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Nutrigénétique de la glutamine et dialogue entre foie et vaisseaux dans l’athérosclérose. La concentration plasmatique de la glutamine (Gln) résulte d’un équilibre entre l’apport alimentaire de Gln, qui augmente sa disponibilité, et sa désamination hépatique par la glutaminase 2 (GLS2), qui convertit Gln en acide glutamique (Glu). Des polymorphismes (single nucleotide polymorphism, SNP) du gène codant GLS2 modulent ce flux métabolique et le rapport Gln/Glu (Gln/Glu ratio, GGR) dans le sang. Ils définissent un nouvel axe nutrigénétique pour contrer une surconsommation de Gln et limiter son accumulation systémique. Un GGR supérieur à 5 est proposé comme un nouveau facteur de risque cardiovasculaire associé à un remodelage matriciel et une inflammation à distance des plaques d’athérosclérose. |
| Dans le texte | |
|
Figure 2 Fluctuation des flux de glutamine et dyspepsie des macrophages dans l’athérosclérose. L’augmentation du flux de glutamine (Gln), illustrée par l’imagerie TEPglutamine, est associée à une augmentation de l’expression du transporteur SLC7A7 par les macrophages de la plaque d’athérosclérose. Cependant, du fait de la diminution de l’expression de la glutaminase 1 (GLS1) dans ces macrophages, les processus anaplérotiques mitochondriaux indispensables à la production d’ATP sont diminués, conduisant à une perte de remodelage des membranes et à un défaut d’efférocytose (« dyspepsie macrophagique »). Le flux de Gln alimente alors préférentiellement la voie de biosynthèse des hexosamines (hexosamine biosynthesis pathway, HBP), ce qui conduit à un remodelage de la matrice extracellulaire (extracellular matrix, ECM). Ces fluctuations métaboliques de la glutamine au sein des macrophages de la plaque d’athérosclérose entraînent sa progression. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.