")
")
Figure 1.
Télécharger l'image originale
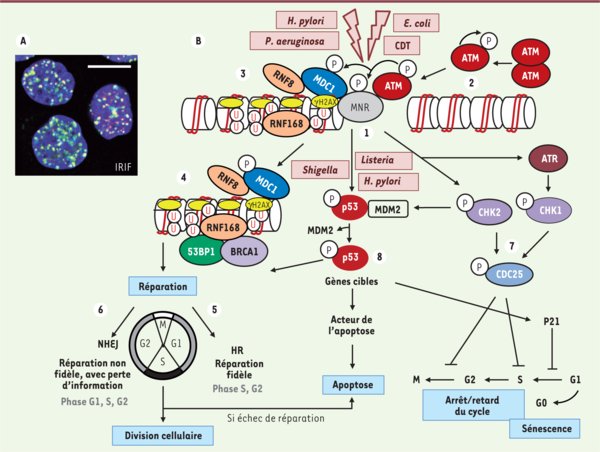
Conséquences des cassures double-brin de l’ADN et mécanismes de réparation. A. Les foyers IRIF marquant les cassures double-brin. Lors d’un stress génotoxique provoquant des cassures double-brin de l’ADN, l’histone H2AX est phosphorylée sur la sérine en position 139 (γH2AX) en quelques minutes. De nombreuses molécules γH2AX s’accumulent sur des centaines de kilobases de part et d’autres des cassures double-brin, formant des foyers nucléaires appelés IRIF (ionizing radiation-induced foci). Chaque foyer témoigne d’une cassure et est constitué d’un macro-complexe de protéines de signalisation et de réparation. En rouge γH2AX, en vert 53BP1, en bleu l’ADN. Le jaune représente la superposition des marquages rouge et vert. Échelle : 10 µm. B. Détection et signalisation des cassures double-brin de l’ADN. Une relaxation de la chromatine est initiée très rapidement au site de cassure de l’ADN (non représentée). Le senseur le plus proximal identifié est le complexe MNR, composé de Mre11, Nbs1 et Rad50, qui possède la capacité intrinsèque de lier l’ADN (étape 1). La kinase ataxia telangiectasia mutated (ATM), inactive sous forme de dimère (étape 2), est activée par autophosphorylation sur sa sérine 1981, et phosphoryle le complexe MNR, H2AX, MDC1 et p53 (étape 3). γH2AX constitue un site de liaison à haute affinité pour la protéine MDC1 qui, à son tour, orchestre le recrutement des autres effecteurs plus tardifs sur la chromatine endommagée. RNF8 et RNF168 sont deux ubiquitine-ligases qui ubiquitinent la chromatine et stimulent la fixation locale des protéines de réparation 53BP1 et BRCA1 (breast cancer 1) (étape 4). Les deux voies de réparation des cassures double-brin sont la recombinaison homologue (HR) et la voie NHEJ (non homologous end joining). La voie HR (étape 5) nécessite une matrice d’ADN dupliqué ; elle est donc active essentiellement à la phase S et G2 du cycle cellulaire. Elle assure une réparation sans erreur et sans perte d’information génétique. La voie NHEJ (étape 6) est active durant tout le cycle cellulaire et peut causer des pertes d’informations génétiques. Le cycle cellulaire est arrêté aux transitions G1/S ou G2/M, après activation de Chk1, Chk2 et Cdc25 (étape 7), donnant le temps aux mécanismes de réparation d’agir afin d’éviter la duplication du génome ou la division cellulaire alors que l’ADN est endommagé. L’apoptose dépendante de p53 intervient quand la cellule ne peut réparer son ADN (étape 8). Les points d’action des différentes bactéries citées dans le texte sont représentés. U : ubiquitine ; P : phosphorylation.
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.